2.1. Общие принципы адаптации на уровне организма
Среда - это все, что окружает организм и прямо или косвенно влияет на его состояние, развитие, рост, выживаемость, размножение и т.д. Составные части и свойства среды многообразны и изменчивы.
Элементы среды, необходимые организму или отрицательно на него воздействующие называются экологическими факторами. По А.С. Мончадскому, экологический фактор – любой элемент или условие среды, на которые организмы реагируют приспособительными реакциями, или адаптациями. Подобное определение исключает такие элементы, как высота над уровнем моря или глубина в водоемах. Действие высоты осуществляется через температуру, инсоляцию (количество лучистой энергии, получаемой поверхностью Земли), атмосферное давление; точно так же глубина влияет на водных обитателей через увеличение давления и уменьшение освещенности.
Экологические факторы принято подразделять на:
- абиотические – это комплекс условий неорганической среды, влияющих на организм;
- биотические – это совокупность влияний жизнедеятельности одних организмов на другие;
- антропогенные – это совокупность воздействия деятельности человека на окружающую среду.
В свою очередь абиотические факторы подразделяются на следующие группы:
- климатические (температура, влажность, ветер и т.д.);
- орогенные (особенности рельефа);
- эдафические (особенности почв);
- гидрологические (соленость, температура и т.д. воды).
Климатические, орогенные и эдафические факторы оказывают преимущественное влияние на наземные организмы, а гидрологические – на гидробионтов.
По другой классификации абиотические факторы подразделяются на физические, химические и эдафические (рис. 28).

Рисунок 28 – Классификация экологических факторов
Один и тот же фактор среды имеет различное значение в жизни совместно обитающих организмов разных видов. Например, сильный ветер зимой неблагоприятен для крупных, обитающих открыто животных, но не действует на более мелких, которые укрываются в норах или под снегом. Солевой состав почвы важен для питания растений, но безразличен для большинства наземных животных и т. п.
Некоторые свойства среды остаются относительно постоянными на протяжении длительных периодов времени в эволюции видов. Таковы сила тяготения, солнечная постоянная, солевой состав океана, свойства атмосферы. Большинство экологических факторов – температура, влажность, ветер, осадки, наличие укрытий, пищи, хищники, паразиты, конкуренты и т. д. – очень изменчиво в пространстве и времени. Степень изменчивости каждого из этих факторов зависит от особенностей среды обитания. Например, температура сильно варьирует на поверхности суши, но почти постоянна на дне океана или в глубине пещер. Паразиты млекопитающих живут в условиях избытка пищи, тогда как для свободноживущих хищников ее запасы все время меняются вслед за изменением численности жертв.
Факторы, изменение которых во времени повторяются регулярно, называются периодическими (приливы и отливы, некоторые океанические течения). Факторы, возникающие неожиданно (извержение вулкана, нападение хищника и т.п.) – непериодические. К ним относятся:
- климатические факторы (ветер, грозы, пожары);
- человеческая деятельность;
- вселение несвойственных данному сообществу хищников, паразитов и патогенных организмов.
Организмы адаптированы к постоянно действующим периодическим факторам, но среди них важно различать первичные и вторичные.
Первичные это те факторы, которые существовали на Земле еще до возникновения жизни: температура, освещенность, приливы, отливы и др.
Вторичные периодические факторы являются следствием изменения первичных: влажность воздуха, зависящая от температуры; растительная пища, зависящая от цикличности в развитии растений. Они возникли позднее первичных и адаптация к ним не всегда четко выражена.
Непериодические факторы обычно воздействуют катастрофически: могут вызвать болезни или даже смерть живого организма.
Среди факторов среды выделяют ресурсы и условия. Ресурсы окружающей среды организмы используют, потребляют, тем самым уменьшая их количество. К ресурсам относят пищу, воду при ее дефиците, убежища, удобные места для размножения и т. п. Условия – это такие факторы, к которым организмы вынуждены приспосабливаться, но повлиять на них обычно не могут. Один и тот же фактор среды может быть ресурсом для одних и условием для других видов. Например, свет –
жизненно необходимый энергетический ресурс для растений, а для обладающих зрением животных – условие зрительной ориентации. Вода для многих организмов может быть и условием жизни, и ресурсом.
Экологические факторы среды оказывают на живые организмы различные воздействия, т. е. могут влиять как раздражители , вызывающие приспособительные изменения физиологических и биохимических функций; как ограничители , обусловливающие невозможность существования в данных условиях; как модификаторы , вызывающие морфологические и анатомические изменения организмов; как сигналы , свидетельствующие об изменениях других факторов среды.
В комплексе действия факторов можно выделить некоторые закономерности, которые являются в значительной мере универсальными (общими) по отношению к организмам.
Правило оптимума. В соответствии с этим правилом для экосистемы, организма или определенной стадии его развития имеется диапазон наиболее благоприятного (оптимального) значения фактора. Для каждого вида растений и животных характерны определенные кардинальные точки воздействия фактора: минимальная, максимальная и оптимальная (рис. 29). За пределами зоны оптимума лежат зоны угнетения, переходящие в критические точки, за которыми существование невозможно. К зоне оптимума обычно приурочена максимальная плотность популяции. Зоны оптимума для различных организмов неодинаковы.
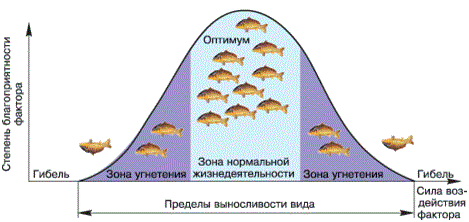
Рисунок 29 – Зависимость результата действия экологического фактора от его интенсивности. Места пересечений кривой с осью абсцисс – критические точки
Свойство видов адаптироваться к тому или иному диапазону факторов среды обозначается понятием экологическая валентность (экологическая пластичность).
Экологически непластичные, то есть маловыносливые виды, организмы с узким диапазоном адаптаций к факторам называются стенобионтными
(греч. stenos – узкий), более выносливые – эврибионтами (греч. eurus – широкий).
Представители разных видов сильно отличаются друг от друга как по положению оптимума, так и по экологической валентности. Так, например, песцы в тундре могут переносить колебания температуры воздуха в диапазоне более 80 0C (от +30 до -5 0C), тогда как тепловодные рачки Copilia mirabilis выдерживают изменения температуры воды в интервале не более 6 0C (от +23 до +29 0C). Одна и та же сила проявления фактора может быть оптимальной для одного вида, пессимальной – для другого и выходить за пределы выносливости для третьего (рис. 30).
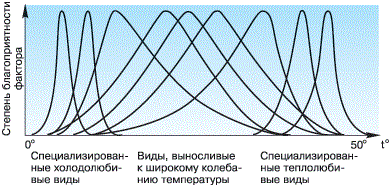
Рисунок 30 – Положение кривых оптимума на температурной шкале для разных видов
Экологическое разнообразие видов проявляется и в положении критических точек: у одних они сближены, у других – широко расставлены. Это значит, что ряд видов может жить только в очень стабильных условиях, при незначительном изменении экологических факторов, а другие выдерживают широкие их колебания. Например, растение недотрога вянет, если воздух не насыщен водяными парами, а ковыль хорошо переносит изменения влажности и не погибает даже в засуху.
Стенобионтность и эврибионтность характеризует различные типы приспособления организмов к выживанию. Виды, длительное время развивающиеся в относительно стабильных условиях, утрачивают экологическую пластичность и вырабатывают черты стенобионтности, в то время как виды, существующие при значительных колебаниях факторов среды, приобретают повышенную экологическую пластичность и становится эврибионтными, т.е. видами с широким диапазоном толерантности.
Условия, приближающиеся по одному или сразу нескольким факторам к критическим точкам, называют экстремальными.
Положение оптимума и критических точек на градиенте фактора может быть в определенных пределах сдвинуто действием условий среды. Это регулярно происходит у многих видов при смене сезонов года. Зимой, например, воробьи выдерживают сильные морозы, а летом гибнут от охлаждения при температуре чуть ниже нуля. Явление сдвига оптимума по отношению к какому–либо фактору носит название акклимации. В отношении температуры это хорошо известный процесс тепловой закалки организма. Для температурной акклимации необходим значительный период времени. Механизмом является смена в клетках ферментов, катализирующих одни и те же реакции, но при разных температурах (так называемые изоферменты ). Каждый фермент кодируется своим геном, следовательно, необходимо выключение одних генов и активация других, транскрипция, трансляция, сборка достаточного количества нового белка и т. п. Общий процесс занимает в среднем около двух недель и стимулируется переменами в окружающей среде. Акклимация, или закалка, – важная адаптация организмов, происходит при постепенно надвигающихся неблагоприятных условиях или при попадании на территории с иным климатом. Она является в этих случаях составной частью общего процесса акклиматизации.
Неоднозначность действия фактора на разные функции.Каждый фактор неодинаково влияет на разные функции организма (рис. 31). Оптимум для одних процессов может являться пессимумом для других. Так, температура воздуха от +40 до +45 0C у холоднокровных животных сильно увеличивает скорость обменных процессов в организме, но тормозит двигательную активность, и животные впадают в тепловое оцепенение. Для многих рыб температура воды, оптимальная для созревания половых продуктов, неблагоприятна для икрометания, которое происходит при другом температурном интервале.
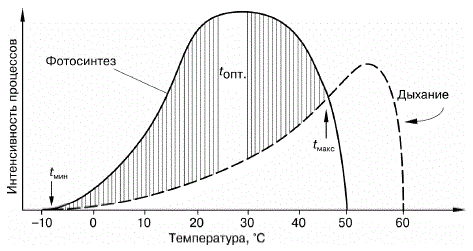
Рисунок 31 – Схема зависимости фотосинтеза и дыхания растения от температуры (по В. Лархеру, 1978): t мин, tопт, tмакс – температурный минимум, оптимум и максимум для прироста растений (заштрихованная область)
Жизненный цикл, в котором в определенные периоды организм осуществляет преимущественно те или иные функции (питание, рост, размножение, расселение и т. п.), всегда согласован с сезонными изменениями комплекса факторов среды. Подвижные организмы могут также менять места обитания для успешного осуществления всех своих жизненных функций.
Степень выносливости, критические точки, оптимальная и пессимальные зоны отдельных индивидуумов не совпадают. Эта изменчивость определяется как наследственными качествами особей, так и половыми, возрастными и физиологическими различиями. Например, у бабочки мельничной огневки – одного из вредителей муки и зерновых продуктов – критическая минимальная температура для гусениц -7 0C, для взрослых форм -22 0C, а для яиц -27 0C. Мороз в -10 0C губит гусениц, но не опасен для имаго и яиц этого вредителя. Следовательно, экологическая валентность вида всегда шире экологической валентности каждой отдельной особи.
Относительная независимость приспособления организмов к разным факторам.Степень выносливости к какому–нибудь фактору не означает соответствующей экологической валентности вида по отношению к остальным факторам. Например, виды, переносящие широкие изменения температуры, совсем не обязательно должны также быть приспособленными к широким колебаниям влажности или солевого режима. Эвритермные виды могут быть стеногалинными, стенобатными или наоборот. Экологические валентности вида по отношению к разным факторам могут быть очень разнообразными. Это создает чрезвычайное многообразие адаптации в природе. Набор экологических валентностей по отношению к разным факторам среды составляет экологический спектр вида.
Несовпадение экологических спектров отдельных видов.Каждый вид специфичен по своим экологическим возможностям. Даже у близких по способам адаптации к среде видов существуют различия в отношении к каким-либо отдельным факторам.
Правило экологической индивидуальности видов сформулировал русский ботаник Л. Г. Раменский (1924) применительно к растениям (рис. 32), затем оно широко было подтверждено и зоологическими исследованиями.
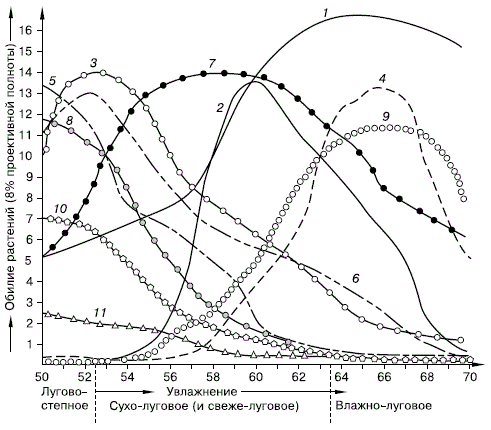
Рисунок 32 – Изменение участия в луговых травостоях отдельных видов растений в зависимости от увлажнения (по Л. Г. Раменскому и др., 1956): 1 – клевер луговой; 2 – тысячелистник обыкновенный; 3 – келерия Делявина; 4 – мятлик луговой; 5 – типчак; 6 – подмаренник настоящий; 7 – осока ранняя; 8 – таволга обыкновенная; 9 – герань холмовая; 10 – короставник полевой; 11 – козлобородник коротконосиковый
Установлено, что для каждого живого организма в отношении различных экологических факторов существуют свои пределы выносливости (толерантности).
Взаимодействие факторов. Все факторы в природе воздействуют на организм одновременно. Причем не в виде простой суммы, а как сложное взаимодействующее соотношение. Такая совокупность факторов называется их констеляцией. Поэтому оптимум и границы выносливости организма по отношению к какому-то одному фактору зависят и от других воздействий. При оптимальной температуре возрастает выносливость к неблагоприятной влажности, недостатку питания. С другой стороны, обилие пищи увеличивает устойчивость организма к изменениям нескольких климатических факторов. Угроза замерзания значительно выше при морозе с сильным ветром, чем в безветренную погоду. Таким образом, один и тот же фактор в сочетании с другими оказывает неодинаковое экологическое воздействие. Наоборот, один и тот же экологический результат может быть получен разными путями. Например, увядание растений можно приостановить путем как увеличения количества влаги в почве, так и снижения температуры воздуха, уменьшающего испарение. Создается эффект частичного взаимозамещения факторов.
Однако эта так называемая «компенсация» факторов ограничена, они не могут заменить друг друга, что, нашло отражение в законе независимости факторов академика В.Р. Вильямса: условия жизни равнозначны, ни один из факторов не может быть полностью заменен другим. Если в количественном выражении хотя бы один из факторов выходит за пределы выносливости вида, то существование последнего становится невозможным, как бы небыли благоприятны остальные условия. Например, нельзя действие влажности заменить действием углекислого газа или солнечного света.
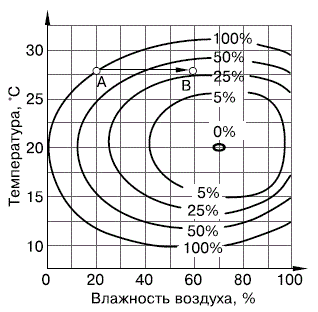
Рисунок 33 – Смертность яиц соснового шелкопряда Dendrolimus pini при разных сочетаниях температуры и влажности
Понятие о влиянии максимального значения фактора, выход, за пределы которого ограничивает жизнедеятельность организмов, ввел в 1911 г. американский зоолог В. Шелфорд на основании изучения размножения жуков–скакунов. Оно отражено в законе толерантности, который иногда называют правилом Шелфорда: присутствие или процветание каких-либо организмов в данном местообитании зависит от комплекса экологических факторов, к каждому из которых у организма существует определенный диапазон толерантности – выносливости. Диапазон устойчивости по каждому фактору ограничен его минимальными и максимальными значениями, в пределах которых только и может существовать организм.
Закон толерантности позднее был дополнен Ю. Одумом (1979) следующими положениями:
- организмы, могут иметь широкий диапазон толерантности в отношении одного экологического и низкий в отношении другого;
- организмы с широким диапазоном толерантности ко всем факторам обычно наиболее широко распространены;
- если условия по одному экологическому фактору не оптимальны для вида, то может сузиться и диапазон толерантности к другим экологическим факторам;
- период размножения является обычно критическим; в этот период многие факторы среды часто становятся лимитирующими (более узкие пределы толерантности для яиц, проростков, личинок и т.д., следовательно, распространение вида часто ограничивается этими стадиями).
Фактор, уровень которого в качественном или количественном отношении (недостаток или избыток) оказывается близким к пределам выносливости данного организма, называется ограничивающим
или лимитирующим. Именно от него и зависит в данный конкретный период выживание особей. В другие отрезки времени ограничивающими могут стать другие факторы, и в течение жизни организмы встречаются с самыми разными ограничениями своей жизнедеятельности (рис. 34). Ограничивающие факторы среды определяют географический ареал вида. Природа этих факторов может быть различной. Так, продвижение вида на север может лимитироваться недостатком тепла, в аридные районы – недостатком влаги или слишком высокими температурами. Лось в Скандинавии встречается значительно севернее, чем в Сибири, хотя в последней среднегодовая температура выше. Причиной препятствующей лосю расширить свой ареал на север в Сибири, оказываются низкие зимние температуры. Рифообразующие кораллы обитают только в тропиках при температуре воды не ниже +20 0С.

Рисунок 34 – Глубина снежного покрова – ограничивающий фактор в распространении оленей
Ограничивающим распространение фактором могут служить и биотические отношения, например занятость территории более сильным конкурентом или недостаток опылителей для растений. Так, опыление инжира всецело зависит от единственного вида насекомых – осы Blastophaga psenes. Родина этого дерева – Средиземноморье. Завезенный в Калифорнию инжир не плодоносил до тех пор, пока туда не завезли ос–опылителей. Распространение бобовых в Арктике ограничивается распределением опыляющих их шмелей. На острове Диксон, где нет шмелей, не встречаются и бобовые, хотя по температурным условиям существование там этих растений еще допустимо.
Чтобы определить, сможет ли вид существовать в данном географическом районе, нужно в первую очередь выяснить, не выходят ли какие–либо факторы среды за пределы его экологической валентности, особенно в наиболее уязвимый период развития.
Выявление ограничивающих факторов очень важно в практике сельского хозяйства, так как, направив основные усилия на их устранение, можно быстро и эффективно повысить урожайность растений или производительность животных. Так, на сильно кислых почвах урожай пшеницы можно несколько увеличить, применяя разные агрономические воздействия, но наилучший эффект будет получен только в результате известкования, которое снимет ограничивающие действия кислотности. Знание ограничивающих факторов, таким образом, ключ к управлению жизнедеятельностью организмов. В разные периоды жизни особей в качестве ограничивающих выступают различные факторы среды, поэтому требуется умелое и постоянное регулирование условий жизни выращиваемых растений и животных.
Впервые на значение лимитирующих факторов указал немецкий ученый Юстус Либих. Еще в 1840 году он установил закон минимума , согласно которому рост растений ограничивается элементом, концентрация которого находится в минимуме. Если в почве полезные компоненты в целом представляют собой уравновешенную систему и только какое-то вещество, например фосфор, содержится в количествах, близких к минимуму, то это может снизить урожай. Но оказалось, что даже те же самые минеральные вещества, очень полезные при оптимальном содержании их в почве, снижают урожай, если они в избытке. Значит, факторы могут быть лимитирующими, находясь и в максимуме.
Совместному действию экологических факторов, а также замещению одних другими посвятил свои исследования швейцарский эколог Е. Рюбель. Он отметил, что каждый экологический фактор только тогда проявляет свое положительное влияние, когда другие факторы не находятся в минимуме. Так, если почва богата питательными веществами, но испытывает недостаток в воде, то растения нормально развиваться не будут (например, при недостатке бора рост растений прекращается, хотя других элементов содержится в избытке).
В. В. Алехин сформулировал близкое к изложенным правило замещения экологических условий : любое условие внешней среды в некоторой степени может замещаться другим; внутренние причины экологических явлений при аналогичном внешнем проявлении могут быть различными.
Например, макроклиматические воздействия могут быть компенсированы биоклиматическими – вечнозеленые виды растут под защитой верхних ярусов растительности, многие виды животных находят микроусловия для существования. Это не агрохимическая и гидробиологическая, а широкоэкологическая и даже биогеографическая трактовка дополнительного правила закона минимума Ю. Либиха.
С законами оптимума и минимума постоянно сталкивается практика сельского хозяйства. Например, рост и развитие пшеницы, а, следовательно, и получение урожая постоянно ограничиваются то критическими температурами, то недостатком или избытком влаги, то нехваткой минеральных удобрений, а иногда и такими катастрофическими воздействиями, как град и бури. Требуется много сил и средств, чтобы поддерживать оптимальные условия для посевов, и при этом в первую очередь компенсировать или смягчать действие именно ограничивающих факторов.
Пути и виды адаптации организмов к условиям окружающей среды.
Адаптация – приспособление организмов к среде; соответствие между условиями окружающей среды и способностью организмов процветать в ней. Этот процесс охватывает строение и функции организмов и их органов. Растения и животные адаптируются к условиям обитания с помощью генетических механизмов, а также посредством более гибких физиологических, поведенческих и эмбриональных механизмов.
В процессе приспособления к неблагоприятным условиям среды организмы смогли выработать три основных пути нейтрализации последних.
Активный путь – это путь, способствующий усилению сопротивляемости, развитию регуляторных процессов, которые позволяют осуществить все жизненные функции организмов, несмотря на факторы. Так, теплокровные животные – птицы и млекопитающие, обитая в условиях изменчивой температуры, поддерживают внутри себя постоянную температуру, которая оптимальна для биохимических процессов в клетках тела. Очевидно, что такое активное сопротивление влиянию внешней среды требует больших затрат энергии, которую им надо постоянно восполнять, а также специальных приспособлений во внешнем и внутреннем строении организмов.
Пассивный путь связан с подчинением жизненных функций организма изменению факторов среды. Так, при недостатке тепла это приводит к угнетению жизнедеятельности и понижению уровня метаболизма, что способствует экономному использованию энергетических запасов.
При резком ухудшении условий среды организмы разных видов могут приостанавливать свою жизнедеятельность и переходить в состояние так называемой скрытой жизни. Например, некоторые мелкие организмы могут полностью высыхать на воздухе, а затем возвращаться к активной жизни после пребывания в воде.
Зимовка (гибернация) – впадение в спячку на зимний период – состояние, подобное сну, в которое впадает животное к началу зимы в основном в умеренных и высоких широтах; при этом все жизненные процессы сведены к минимуму. Так, зимняя спячка у млекопитающих (сурок, бурый медведь, летучие мыши, ежи и др.) длится с осени до весны (6–8 месяцев) (рис. 35). Эти животные способны понижать температуру своего тела почти до нуля, замедляя метаболизм и, тем самым, трату питательных веществ. Во время спячки температура тела падает до 3-4 0
С, число сердечных сокращений – до 4-20 (в активном состоянии – 500-700), число дыхательных движений – до 2-3 (вместо 100-150) и т.д. Во время спячки у животных отмечается особый вид пищевого поведения: длительное голодание при отсутствии чувства голода, существование механизма поддержания веса тела. Все системы организма во время гибернации работают четко и согласованно. Пробуждение из спячки – кратковременный стремительный процесс: за три часа температура тела животного повышается до 37 0С, потребление кислорода возрастает в 100-150 раз. Из безжизненного животное быстро превращается в активное, используя лишь собственные ресурсы.
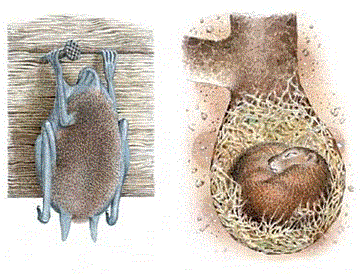
Рисунок 35 – Летучая мышь ушан и суслик в состоянии зимней спячки
Спячка летняя (эстивация) – неактивное состояние, подобное сну (летний покой) некоторых животных (грызунов) в аридных районах, помогающее им пережить сухой и жаркий период года. Сюда также можно отнести летнее оцепенение пойкилотермных животных (клещей, некоторых насекомых и др.).
Во временное оцепенение могут впадать некоторые виды птиц (рис. 36). Так стриж – насекомоядная птица, добывающая корм в полете. Интенсивный обмен веществ определяет большие энергетические потребности организма этой небольшой птицы. Ночью или во время непогоды стриж не питается, поэтому впадает в оцепенение (температура тела падает, замедляется скорость обменных процессов).

Рисунок 36 – Оцепенение птенцов стрижа черного (Apus apus)
Анабиоз – способность организмов переживать неблагоприятное время (изменение температуры окружающей среды, отсутствие влаги и др.) в состоянии, при котором резко снижается обмен веществ, и отсутствуют видимые проявления жизни. При наступлении благоприятных условий организм оживает. Явление анабиоза впервые было обнаружено А. Левенгуком (1701). Наблюдаемые им «анималькули»
(коловратки) при высыхании капли воды сморщивались, выглядели мертвыми и могли пребывать в таком состоянии длительное время. Помещенные вновь в воду, они набухали и переходили к активной жизни. Левенгук объяснил это явление тем, что оболочка «анималькулей », очевидно, «не позволяет ни малейшего испарения» и они остаются живыми в сухих условиях. Однако через несколько десятилетий естествоиспытатели уже спорили о возможности того, что « жизнь может быть полностью прекращена» и восстановлена вновь через 20, 40, 100 лет или более ».
В 70– х годах XVIII в. явление «воскрешения» после высыхания было обнаружено и подтверждено многочисленными опытами у ряда других мелких организмов – пшеничных угриц, свободноживущих нематод и тихоходок. Ж. Бюффон, повторив опыты Дж. Нидгема с угрицами, утверждал, что «эти организмы можно заставить сколько угодно раз подряд умирать и вновь оживать» Л. Спалланцани впервые обратил внимание на глубокий покой семян и спор растений, расценив его как сохранение их во времени.
В середине XIX в. было убедительно установлено, что устойчивость сухих коловраток, тихоходок и нематод к высоким и низким температурам, недостатку или отсутствию кислорода возрастает пропорционально степени их обезвоживания. Однако оставался открытым вопрос, происходит ли при этом полное прерывание жизни или лишь ее глубокое угнетение. В 1878 г. Клод Бернал выдвинул понятие «скрытая жизнь», которую он характеризовал прекращением обмена веществ и «перерывом отношений между существом и средой».
Окончательно этот вопрос был решен лишь в первой трети XX столетия с развитием техники глубокого вакуумного обезвоживания. Опыты Г. Рама, П. Беккереля и других ученых показали возможность полной обратимой остановки жизни. В сухом состоянии, когда в клетках оставалось не более 2 % воды в химически связанном виде, такие организмы, как коловратки, тихоходки, мелкие нематоды, семена и споры растений, споры бактерий и грибов выдерживали пребывание в жидком кислороде (– 218,4 0C), жидком водороде (– 259,4 0C), жидком гелии (– 269,0 0C), т. е. температуры, близкие к абсолютному нулю. При этом содержимое клеток затвердевает, отсутствует даже тепловое движение молекул, и всякий обмен веществ, естественно, прекращен. После помещения в нормальные условия эти организмы продолжают развитие. У некоторых видов остановка обмена веществ при сверхнизких температурах возможна и без высушивания, при условии замерзания воды не в кристаллическом, а в аморфном состоянии.
Полная временная остановка жизни получила название анабиоза. Термин был предложен В. Прейером еще в 1891 г. В состоянии анабиоза организмы становятся устойчивыми к самым разнообразным воздействиям. Например, тихоходки выдерживали в эксперименте ионизирующее облучение до 570 тыс. рентген в течение 24 ч. Обезвоженные личинки одного из африканских комаров-хирономусов – Polypodium vanderplanki – сохраняют способность оживать после воздействия температуры в +102 0C.
Состояние анабиоза намного расширяет границы сохранения жизни, в том числе и во времени. Например, в толще ледника Антарктиды при глубоком бурении были обнаружены микроорганизмы (споры бактерий, грибов и дрожжей), развившиеся впоследствии на обычных питательных средах. Возраст соответствующих горизонтов льда достигает 10-13 тыс. лет. Споры некоторых жизнеспособных бактерий выделены и из более глубоких слоев возрастом в сотни тысяч лет. Скрытая жизнь семян многих растений, сохраняющих всхожесть порой десятки лет. Так, сухие семена могут долго лежать, не теряя всхожести: семена мака – до 10 лет, зерновки ржи, ячменя и пшеницы – до 32, плоды одуванчика – до 68, лотоса – до 250 лет. Известен случай, когда проросли семена лотоса, найденные в торфе болота, высохшего 2000 лет тому назад. Плоды этого растения покрыты толстой газо– и водонепроницаемой оболочкой.
Анабиоз, однако, – достаточно редкое явление. Он возможен далеко не для всех видов и является крайним состоянием покоя в живой природе. Его необходимое условие – сохранение неповрежденными тонких внутриклеточных структур (органелл и мембран) при высушивании или глубоком охлаждении организмов. Это условие невыполнимо для большинства видов, имеющих сложную организацию клеток, тканей и органов.
Способность к анабиозу обнаруживается у видов, имеющих простое или упрощенное строение и обитающих в условиях резкого колебания влажности (пересыхающие мелкие водоемы, верхние слои почвы, подушки мхов и лишайников и т. п.).
Гораздо шире распространены в природе другие формы покоя, связанные с состоянием пониженной жизнедеятельности в результате частичного угнетения метаболизма. Любая степень снижения уровня обмена веществ повышает устойчивость организмов и позволяет более экономно тратить энергию.
Формы покоя в состоянии пониженной жизнедеятельности делят на гипобиоз и криптобиоз, или покой вынужденный
и покой физиологический. При гипобиозе торможение активности, или оцепенение, возникает под прямым давлением неблагоприятных условий и прекращается почти сразу после того, как эти условия возвращаются к норме (рис. 37). Подобное подавление процессов жизнедеятельности может возникать при недостатке тепла, воды, кислорода, при повышении осмотического давления и т. п. В соответствии с ведущим внешним фактором вынужденного покоя различают криобиоз (при низких температурах), ангидробиоз (при недостатке воды), аноксибиоз (в анаэробных условиях), гиперосмобиоз (при высоком содержании солей в воде) и др.
He только в арктических и антарктических, но и в средних широтах некоторые морозостойкие виды членистоногих (коллемболы, ряд мух, жужелицы и др.) зимуют в состоянии оцепенения, быстро оттаивая и переходя к активности под лучами солнца, а затем вновь теряют подвижность при снижении температуры. Взошедшие весной растения прекращают и возобновляют рост и развитие вслед за похолоданием и потеплением. После выпавшего дождя голый грунт часто зеленеет за счет быстрого размножения почвенных водорослей, находившихся в вынужденном покое.
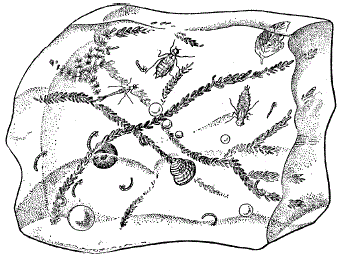
Рисунок 37 – Пагон – кусок льда со вмерзшими в него пресноводными обитателями (из С. А. Зернова, 1949)
Глубина и продолжительность подавления обмена веществ при гипобиозе зависит от длительности и интенсивности действия угнетающего фактора. Вынужденный покой наступает на любой стадии онтогенеза. Выгоды гипобиоза – быстрое восстановление активной жизнедеятельности. Однако это относительно неустойчивое состояние организмов и при большой длительности может быть повреждающим из-за разбалансированности метаболических процессов, истощения энергетических ресурсов, накопления недоокисленных продуктов обмена и других неблагоприятных физиологических изменений.
Криптобиоз – принципиально другой тип покоя. Он связан с комплексом эндогенных физиологических перестроек, которые происходят заблаговременно, до наступления неблагоприятных сезонных изменений, и организмы оказываются к ним готовы. Криптобиоз является адаптацией прежде всего к сезонной или иной периодичности абиотических факторов внешней среды, их регулярной цикличности. Он составляет часть жизненного цикла организмов, возникает не на любой, а на определенной стадии индивидуального развития, приуроченной к переживанию критических периодов года.
Переход в состояние физиологического покоя требует времени. Ему предшествует накопление резервных веществ, частичная дегидратация тканей и органов, уменьшение интенсивности окислительных процессов и ряд других изменений, понижающих в целом тканевый метаболизм. В состоянии криптобиоза организмы становятся во много раз более устойчивыми к неблагоприятным воздействиям внешней среды (рис. 38). Основные биохимические перестройки при этом являются во многом общими для растений, животных и микроорганизмов (например, переключение метаболизма в разной степени на путь гликолиза за счет резервных углеводов и т. п.). Выход из криптобиоза также требует времени и затрат энергии и не может быть осуществлен простым прекращением отрицательного действия фактора. Для этого необходимы особые условия, различные для разных видов (например, промораживание, присутствие капельно-жидкой воды, определенная продолжительность светового дня, определенное качество света, обязательные колебания температуры и др.).
Криптобиоз как стратегия выживания в периодически неблагоприятных для активной жизни условиях – это продукт длительной эволюции и естественного отбора. Он широко распространен в живой природе. Состояние криптобиоза характерно, например, для семян растений, цист и спор различных микроорганизмов, грибов, водорослей. Диапауза членистоногих, спячка млекопитающих, глубокий покой растений – также различные типы криптобиоза.
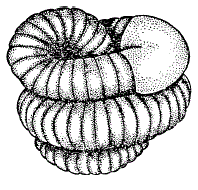
Рисунок 38 – Дождевой червь в состоянии диапаузы (по В. Тишлеру, 1971)
Состояния гипобиоза, криптобиоза и анабиоза обеспечивают выживание видов в природных условиях разных широт, часто экстремальных, позволяют сохранять организмы в течение длительных неблагоприятных периодов, расселяться в пространстве и во многом раздвигают границы возможности и распространения жизни в целом.
Кроме подчинения и сопротивления воздействию внешней среды, возможен и третий способ выживания – избегание неблагоприятных условий и активный поиск других, более благоприятных местообитаний.
Обычный для животных путь приспособления к неблагоприятным периодам – миграция (рис. 39). Так, сайгаки ежегодно уходят на зиму в малоснежные южные полупустыни, где зимние травы в связи с сухостью климата более питательны и доступны. Однако летом травостои полупустынь быстро выгорают, поэтому на период размножения сайгаки переходят в более влажные северные степи.
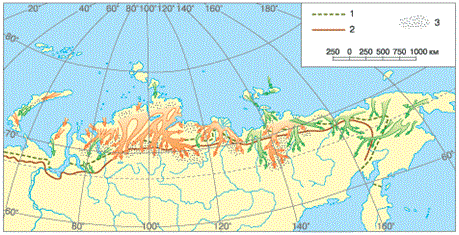
Рисунок 39 – Кочёвки северных оленей: 1 – северная граница лесотундры; 2 – северная граница тайги; 3 – места зимовок
Зимующие тетерева и рябчики на большую часть суток зарываются в снег, где гораздо теплее. Многие животные устраивают жилища – норы и гнезда, защищающие их от внешних воздействий. Это тоже путь избегания неблагоприятных факторов (рис. 43).
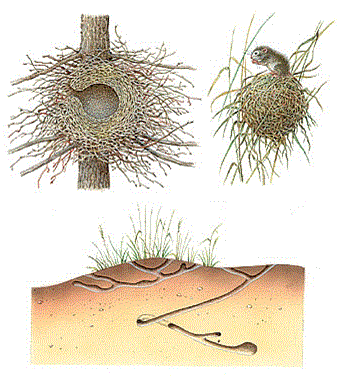
Рисунок 40 – Гнезда и норы животных. Вверху: слева – гнездо обыкновенной белки; справа – гнездо мыши–малютки. Внизу летняя (слева) и зимняя (справа) норы полуденных песчанок
Ярким примером избегания зимней бескормицы и холодов являются дальние перелеты птиц (рис. 41).
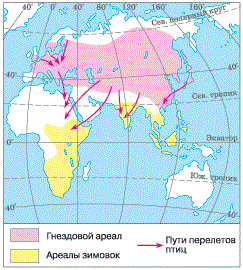

Рисунок 41 – Карта миграции: слева – миграция деревенской ласточки; справа – миграция журавлей
Все три пути выживания могут сочетаться у представителей одного и того же вида. Например, растения не могут поддерживать постоянную температуру тела, но многие из них способны регулировать водный обмен. Холоднокровные животные подчиняются неблагоприятным факторам, но могут и избежать их воздействия. В целом, при огромном разнообразии живой природы в ней можно выделить лишь несколько основных путей приспособительного развития видов.
Основные механизмы адаптации на уровне организма. К основным механизмам приспособления к среде на уровне организма относятся морфологические, физиологические поведенческие адаптации.
Морфологические адаптации выражаются в наличии таких особенностей внешнего строения, которые способствуют выживанию и успешной жизнедеятельности организмов в обычных для них условиях. Это, в частности, приспособления к скоростному плаванию у китообразных, к парению в воде у планктонных организмов. Растения, обитающие в пустыне, лишены листьев, и их строение наилучшим образом приспособлено к минимальным потерям влаги. Так, для некоторых рыб мелководья яркий свет становиться средством защиты – благодаря их прозрачности. Хотя скелет и внутренние органы стеклянного сома светонепроницаемы, тело рыбы почти все прозрачно. Поэтому хищник, прямо смотрящий на сома, при обычных обстоятельствах увидит лишь испещренные солнечными пятнами подводные заросли позади него. Некоторые насекомые в ходе эволюции также стали прозрачными.
Приспособительная окраска является одним из важных средств пассивной защиты организмов. Многие животные средних и высоких широт зимой имеют белую окраску (песец, заяц, горностай). Броские полосы тигра, создающие так называемую расчленяющую окраску, представляют собой форму маскировки, которая помогает огромной кошке во время охоты затаиться в высоких травах. Среди саранчовых насекомых обитатели густой злаковой растительности отличаются зеленым цветом, стройным, сжатым с боков телом с гладкими покровами, заостренной формой головы. У обитателей открытых пустынных участков тело коренастое, широкое, покрыто бугорками и морщинками, окрашено под цвет грунта, угол головы тупой, задние бедра очень мощные (рис. 42).

Рисунок 42 – Два вида саранчи: обитатель травостоя (слева) и каменистого грунта (справа)
Морфологический тип приспособления животного или растения, при котором они имеют внешнюю форму, отражающую способ взаимодействия со средой обитания, называют жизненной формой вида. При этом разные виды будут иметь сходную жизненную форму, если ведут близкий образ жизни. Примерами здесь могут быть кит (млекопитающее), пингвин (птица) и акула (рыба).
Физиологические адаптации проявляются, например, в особенностях ферментативного набора в пищеварительном тракте животных, определяемого составом пищи. Например, борьба с перегревом путем увеличения испарения, борьба с охлаждением у пойкилотермных животных путем частичного обезвоживания своего тела или накопления специальных веществ, понижающих точку замерзания, у гомойотермных – за счет изменения обмена веществ. Так, верблюд способен обеспечивать потребности во влаге путем биохимического окисления собственного жира.
Поведенческие адаптации проявляются в самых различных формах. Примерами могут служить формы приспособительного поведения животных, направленные на обеспечение нормального теплообмена с окружающей средой: создание убежищ, передвижение с целью выбора оптимальных температурных условий, особенно в условиях экстремальных (очень высоких или очень низких) температур. Известны суточные и сезонные кочевки млекопитающих и птиц. Приспособительные поведение может проявляться у хищников в процессе выслеживания и преследования добычи, а жертв – в определенных ответных реакциях (например, затаивание). Некоторые насекомые отпугивают хищников и паразитов резкими движениями.
Мимикрия – это подражание животных или растений определенным предметам неживой или живой природы. Так, морской дракон, обитающий в водах у берегов Австралии, имеет совершенную маскировку. Плавники в форме листьев делают его похожим на плывущее растение; и он даже двигает ими, как колыхающееся в струе течения водное растение. В 1862 г. английский натуралист Г. Бейтс, изучая животных в бассейне р. Амазонка, установил подражание незащищенного съедобного вида (имитатора) другому, неродственному ему виду, который невкусен или опасен для хищника. Так, многие виды безвредных змей сходны по внешнему виду с ядовитыми; безвредными мухи и бабочки-стеклянницы часто подражают пчелам и осам и др., то есть это один из типов покровительственной окраски у животных (мимикрия Бейтса).
Мимикрия Мюллера. Этот вид мимикрии был открыт немецким зоологом Ф. Мюллером в 1878 г. при изучении насекомых в Бразилии. Несколько защищенных видов животных имеют сходную внешность и, подражая друг другу по окраске и форме, образуют кольцо. Например, многие виды ос сходны по очертаниям тела и окраске, ядовитые насекомые (семиточечная божья коровка, клоп-солдатик, жук-нарывник) имеют красную окраску с черными пятнами (отпугивающая окраска). Враги насекомых, выработав рефлекс отвращения на один вид, уже не трогают насекомых др. видов, входящих в это кольцо. Эффект мимикрии усиливается особенностями поведения животных. Так, у многих видов пчел и ос на теле имеются желтые и черные полоски, и хищники, часто сталкиваясь с некоторыми видами–имитаторами, привыкают избегать их, что выгодно всем видам, входящим в кольцо мимикрии. Жуки–дровосеки могут подражать осам, муравьями и выглядеть как несъедобные жуки из других групп. Такая защитная окраска весьма необходима для лучшего выживания организмов.
Мимикрия у растений – служит для привлечения или отпугивания животных. Это обычно касается отдельных организмов, а не организма в целом. Растения обманывают животных, подражая другим растениям – моделям. Так, лишенные нектара цветки внешне сходны с медоносными, привлекают насекомых, которые в поисках нектара опыляют их. Ловчие аппараты насекомоядных растений (непентес) часто напоминают яркие цветки других растений, привлекая этим насекомых, которые погибают в ловушках.
Принципы экологической классификации организмов. Экологические классификации отражают сходство, возникающее у представителей самых разных групп, если они используют сходные пути адаптации. Например, если мы классифицируем животных по способам движения, то в экологическую группу видов, передвигающихся в воде реактивным путем, попадут такие разные по систематическому положению животные, как медузы, головоногие моллюски, некоторые инфузории и жгутиковые, личинки ряда стрекоз и др. (рис. 43).
В основе экологической классификации может быть положены и места обитания. Водные организмы при этом подразделяются на бентосные, планктонные и нектонные.
Разделение всех организмов на эврибионтных и стенобионтных по широте диапазона приспособлений к среде представляет пример простейшей экологической классификации.
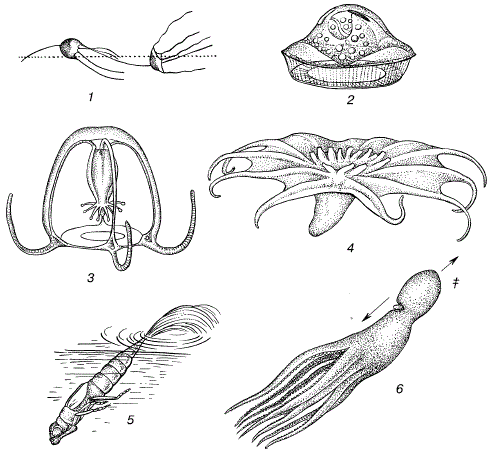
Рисунок 43 – Представители экологической группы организмов, передвигающихся в воде реактивным способом (по С. A. Зернову, 1949):
- 1 – жгутиковое Medusochloris phiale;
- 2 – инфузория Craspedotella pileosus;
- 3 – медуза Cytaeis vulgaris;
- 4 – пелагическая голотурия Pelagothuria;
- 5 – личинка стрекозыа–коромысла;
- 6 – плывущий осьминог Octopus vulgaris:
а–направление струи воды;
б–направление движения животного
Другой пример – разделение организмов на группы по характеру питания. Автотрофы – это организмы, использующие в качестве источника для построения своего тела неорганические соединения. Гетеротрофы – все живые существа, нуждающиеся в пище органического происхождения. В свою очередь, автотрофы делятся на фототрофов и хемотрофов . Первые для синтеза органических молекул используют энергию солнечного света, вторые – энергию химических связей. Гетеротрофов делят на сапрофитов , использующих растворы простых органических соединений, и голозоев. Голозои обладают сложным комплексом пищеварительных ферментов и могут употреблять в пищу сложные органические соединения, разлагая их на более простые составные компоненты. Голозои делятся на сапрофагов (питаются мертвыми растительными остатками), фитофагов (потребителей живых растений), зоофагов (нуждающихся в живой пище) и некрофагов (трупоядных животных). В свою очередь, каждую из этих групп можно подразделить на более мелкие, имеющие свою специфику в характере питания.
Иначе можно построить классификацию по способу добывания пищи. Среди животных выявляются, например, такие группы, как филътраторы (мелкие рачки, беззубка, кит и др.), пасущиеся формы (копытные, жуки-листоеды), собиратели (дятлы, кроты, землеройки, куриные), охотники на движущуюся добычу (волки, львы, мухи–ктыри и т. п.) и целый ряд других групп. Так, несмотря на большое несходство в организации, одинаковый способ овладения добычей приводит у львов и мух–ктырей к ряду аналогий в их охотничьих повадках и общих чертах строения: поджарости тела, сильному развитию мускулатуры, способности развивать кратковременно большую скорость и т. п.
Наиболее распространена экологическая классификация организмов по жизненным формам. Что это такое жизненная форма? В трактовке И.Г. Серебрякова(1964), « жизненная форма – это своеобразная внешняя форма организмов, обусловленная биологией развития и внутренней структурой их органов, формируется в определенных почвенно-климатических условиях, как приспособление жизни к этим условиям», то есть это форма организмов, приспособившихся к своей среде обитания под длительным влиянием комплекса факторов. Его же определение, но более короткое: « Жизненная форма растения – это его габитус (внешняя форма вида), связанный с ритмом развития и приспособленный к современным и прошлым условиям среды».
Изучение жизненных форм, их динамики исключительно важно для познания закономерностей приспособления популяций и организмов к тем или иным условиям среды. Сообщества растений и животных устойчивее и продуктивнее, если они состоят из видов с разными жизненными формами. Такое сообщество полнее использует ресурсы среды и имеет более разнообразные внутренние связи. Без изучения жизненных форм невозможно изучить ценотическую структуру – один из основных аспектов структурно-функциональной организации биогеоценоза.
Классификации жизненных форм животных. Наиболее известны классификации Д.Н. Кашкарова (1945). По приспособлению к передвижению Д.Н. Кашкаров выделяет следующие жизненные формы животных.
Плавающие формы.
- Чисто водные: а) нектон, б) планктон, в) бентос.
- Полуводные: а) ныряющие, б) неныряющие, в) добывающие из воды лишь пищу.
Роющие формы.
- Абсолютные землерои (всю жизнь проводят под землей).
- Относительные землерои (выходят на поверхность земли).
Наземные формы.
- Не делающие нор: а) бегающие, б) прыгающие, в) ползающие.
- Делающие норы: а) бегающие, б) прыгающие, в) ползающие.
- Животные скал.
Древесные лазающие формы: а) не сходящие с деревьев; б) лишь лазающие по деревьям.
Воздушные формы: а) добывающие пищу в воздухе; б) высматривающие ее с воздуха.
По отношению к влажности воздуха Кашкаров выделяет влаголюбивые (гигрофильные) и сухолюбивые (ксерофильные) формы; по питанию – растительноядных, всеядных, хищных, могильщиков (трупоедов); по месту размножения – размножающихся под землей, на поверхности земли, в ярусе трав, в кустарниках, на деревьях.
Разные категории жизненных форм насекомых (геобионты, гидробионты и т.д.) относительно среды обитания разработали В.В. Яхонтов, Ф.Н. Правдин, И.Х. Шарова.
Классификации жизненных форм растений. Первая классификация жизненных форм принадлежит древнегреческому ученому и философу Теофрасту. Он разделил все растения на деревья, кустарники, полукустарники и травы.
Основоположник биогеографии А. Гумбольдт в 1806 году по приспособлению к комплексу факторов выделил 19 «основных форм » растений, в том числе: 1) форма пальм; 2) форма бананов; 3) форма хвойных деревьев; 4) форма кактусовидных растений; 5) форма лиан; 6) форма лавровых деревьев; 7) форма злаковидная; 8) форма мхов; 9) форма орхидей;
Из множества классификаций жизненных форм растений наибольшее признание получили две: К. Раункиера (1906) и И.Г. Серебрякова (1964).
За основу классификации К. Раункиера принято (рис. 44) расположение почек возобновления и наличие приспособлений для переживания неблагоприятного сезона года (в умеренных и арктических широтах – зима, в аридных районах еще и летние засухи). Выбранный ученым признак коррелятивно связан со множеством других. Все растения К. Раункиер разделил на 5 типов жизненных форм, которые называл также биологическими типами. Каждый из типов жизненных форм Раункиера подразделяется еще на 3-15 подтипов, характеризующихся разным характером и расположением побегов, защищенностью почек и другими морфологическими признаками.
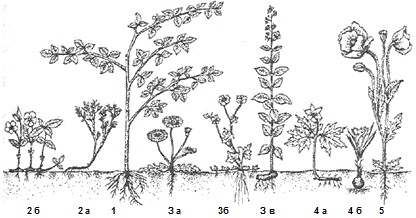
Рисунок 44 – Жизненные формы установленные Раункером (по Г.И. Поплавской, 1948): 1 – фанерофиты (береза); 2 – хамефиты (гвоздика травянка, брусника); 3 – гемикриптофиты (одуванчик, лютик, вербейник); 4 – геофиты (ветреница, тюльпан); 5 – терофиты (мак)
Фанерофиты (Ph) – почки возобновления, открытые или закрытые, расположены на вертикально растущих побегах высоко над землей (выше 30 см). Деревья, кустарники, лианы, стеблевые суккуленты и стеблевые травы. Эта жизненная форма подразделена на 15 подтипов.
- Травянистые фанерофиты растут в условиях постоянно влажного тропического климата. Они напоминают высокие травы умеренного пояса, но их побеги живут несколько лет, не деревенея, а стебли, как правило, слабее, чем у деревянистых растений. К ним относятся бегония и многие виды семейств крапивных, бальзаминовых, молочайных, перечных, ароидных, коммелиновых н др.
- Вечнозеленые мегафанерофиты – растения более 30 м высоты с незащищенными почками.
- Вечнозеленые мезофанерофиты – растения высотой 8-30 м с незащищенными почками.
- Вечнозеленые микрофанерофиты – растения высотой 2-8 м с незащищенными почками.
- Вечнозеленые нанофанерофиты – растения ниже 2 м с незащищенными почками.
2-5-я группы объединяют деревянистые растения влажных тропических лесов.
- Эпифитные фанерофиты – цветковые и папоротникообразные эпифиты тропических и субтропических лесов.
- Вечнозеленые мегафанерофиты с защищенными почками.
- Вечнозеленые мезофанерофиты с защищенными почками.
- Вечнозеленые микрофанерофиты с защищенными почками.
- Вечнозеленые нанофанерофиты с защищенными почками.
7-10-я группы объединяют деревянистые растения субтропических лавровых и жестколистных лесов, а также хвойные деревья и кустарники. К 10-й группе относят также вечнозеленые кустарнички умеренных и холодных широт (брусника, толокнянка и др.).
- Стеблесуккулентные фанерофиты – кактусы, кактусовидные молочаи и др.
- Мегафанерофиты с опадающей листвой и защищенными почками.
- Мезофанерофиты с опадающей листвой и защищенными почками.
- Микрофанерофиты с опадающей листвой и защищенными почками.
К 12-14-й группам автор относит деревья и кустарники лесов с опадающей на сухое или на холодное время года листвой, а также деревянистые растения саванн.
- Нанофанерофиты с опадающей листвой и защищенными почками. Кустарнички умеренных и холодных широт с опадающей на зиму листвой (черника, карликовая березка и др.).
Хамефиты (Ch) – почки возобновления близко к поверхности, не выше 20-30 см. В умеренных широтах побеги этих растений зимуют под снегом и не отмирают. Травянистые растения, кустарнички (черника, линнея северная, брусника, багульник простертый, морошка, дерен канадский).
Эту жизненную форму автор подразделяет на четыре подтипа:
- Полукустарниковые хамефиты, верхние части побегов которых отмирают к концу вегетационного периода, так что только нижние их части переносят неблагоприятный период. Виды этого подтипа произошли частично от травянистых фанерофитов, частично – от нанофанерофитов. Особенно характерны они для средиземноморского климата. К ним относятся виды семейств губоцветных, гвоздичных, бобовых и др. К этому же подтипу относятся и хамефиты с поднимающимися кверху побегами, не отмирающими на концах, но имеющими ограниченный рост.
- Пассивные хамефиты, вегетативные побеги которых отрицательно геотропичны и остаются неизменными в начале неблагоприятного периода. Они слабы, у них нет достаточно развитой механической ткани и поэтому не могут стоять прямо, падают и лежат на земле. На концах они приподнимаются, так как рост концов побегов вызывает отрицательный геотропизм. Этот подтип включает виды вечнозеленые и с опадающей листвой, с защитой почек и без защиты. Одни из этих растений травянистые, другие деревянистые. Особенно их много в альпийской области гор. Сюда относятся виды резухи (Arabis), очитка (Sedum), камнеломки (Saxifraga), крупки (Draba), а из растений равнин звездчатка жестколистная (Stellaria holostea) и др.
- Активные хамефиты, вегетативные побеги которых остаются неизмененными в начале неблагоприятного периода. Эти побеги лежат на поверхности земли потому, что они трансверсально (поперечно) геотропичны. Поэтому в отличие от побегов пассивных хамефитов побеги этих растений на концах не приподнимаются. В этом подтипе, как и в предыдущем, объединены виды вечнозеленые и с опадающей листвой, с защитой почек и без защиты, травянистые и деревянистые. Сюда относятся виды барвинка (Viпca), тимьяна (Thyтus), а также вероника лекарственная (Veroпica officiпalis), толокнянка (Arctostaphylos), линнея северная (ипnаеа borealis), вороника черная (Eтpetruт nigrum), луговой чай (Lysiтachia пuттularia) и др.
- Растения–подушки. Побеги их отрицательно геотропичны, как и у пассивных хамефитов, но растут они так тесно, что не дают упасть друг другу, даже если механическая ткань развита слабо. Побеги короткие. Подушковидный рост защищает от неблагоприятных условий среды. Эта группа произошла от пассивных хамефитов. Она еще в большей степени, чем группа пассивных хамефитов, характерна для альпийской области гор. К нейотносятся некоторые высокогорные виды незабудок (Myosotis), камнеломок (Saxifraga), соссюрей (Saussurea) и др.
Гемикриптофиты (Hk) – почки возобновления на поверхности почвы или в самом поверхностом слое, под подстилкой. Дернообразующие, надземные побеги к зиме отмирают. Многие луговые и лесные растения (одуванчик, злаки, осоки, лютик Франше, лапчатка земляниколистная, хохлатка бледная, пионы, башмачки, крапива).
Эту жизненную форму автор подразделяет на три подтипа:
- Протогемикриптофиты. К этому подтипу относятся гемикриптофиты, у которых воздушные побеги, несущие листья и цветки, удалены от основания. Наиболее крупные листья находятся в средней части побега, и их размеры уменьшаются книзу и кверху от средней части. Книзу листья становятся чешуйчатыми и служат для защиты почек в неблагоприятный период. Они ежегодно образуют не цветущие удлиненные воздушные побеги, которые при благоприятных условиях могут пережить зиму, и в этом случае растение ведет себя как полукустарниковый хамефит.
Этот подтип встречается там, где неблагоприятный период вызван либо засухой, либо холодом.
У одних протогемикриптофитов отсутствуют столоны (зверобои – Hypericum, молочаи – Euphorbia, норичники – Scrophularia и др.), другие снабжены подземными или надземными столонами (кипрей болотный – Epilobium palustre, чистец лесной – Stachys sylvatica, крапива двудомная – Urtica dioica, василистник малый – Thalictrum miпus, некоторые льнянки – Linaria).
Столонами называют подземные или надземные стелющиеся по поверхности почвы недолговечные побеги с длинными междоузлиями, служащие для размножения.
Виды рода малина (Rubus) характеризуются тем, что они в первый год дают вегетативные побеги, которые, перезимовав, развивают боковые цветущие ветви. После плодоношения побеги отмирают. Таким образом, на воздушной части вегетативных побегов находятся только почки, из которых развиваются цветущие побеги, а вегетативные почки, от которых зависит продолжение индивидуальной жизни растения, располагаются на подземной части побега. Это дает основание причислить род малина (к их числу относится малина обыкновенная – Rubus idaeus) к протогемикриптофитам.
- Частично розеточные гемикриптофиты. К этому подтипу автор относит гемикриптофиты, у которых воздушные побеги, несущие и листья и цветки, характеризуются тем, что самые крупные листья и обычно в наибольшем числе находятся в нижней части побега, где междоузлия более или менее укорочены, так что листья образуют род розетки. Эти растения обитают преимущественно в умеренном климате, где лето не очень сухое и где почва покрыта снегом на более или менее длительный срок.
Кроме многолетних растений, к этой группе отнесены и очень многие двулетние. Подавляющее большинство растений этой группы не образует столонов (таковы многие гвоздичные, лютиковые, розоцветные, зонтичные, колокольчиковые, сложноцветные, дерновинные злаки и другие растения). У некоторых имеются воздушные (живучка ползучая – Ajuga repiaпs) или подземные (сныть обыкновенная – Aegopodium podagraria) столоны.
- Розеточные гемикриптофиты. К этому подтипу относятся гемикриптофиты, у которых удлиненная надземная часть побега несет только цветки, а листья сосредоточены у основания побега. В большинстве случаев эти растения в первый год развивают розетку листьев и лишь на второй год дают безлистный вертикальный надземный побег. Они обитают преимущественно в районах со снежным покровом. Многие из них имеют вечнозеленые листья. Из розеточных гемикриптофитов не образуют столонов росянка (Drosera), кермек (Statice), первоцвет (Primula), маргаритка (Bellis), одуванчик (Taraxacum), кульбаба (Leoпtodoп) и др. Имеет столоны подбел (Petasites).
Протогемикриптофиты и розеточные гемикриптофиты представляют собой два резко различных типа растений: пластичный и консервативный. Протогемикриптофиты – пластичные растения. У них к осени развивается большое количество облиственных побегов различной длины и почек, расположенных как у корневой шейки, так и выше и ниже ее. В зависимости от суровости зимы они сохраняют то только почки, то почки и побеги с листьями, находящиеся на разных стадиях развития и расположенные на различной высоте над корневой шейкой. Они приспособлены к меняющимся год от года метеорологическим особенностям зимы, но в суровые зимы теряют часть своих ростков и побегов. Розеточные гемикриптофиты – консервативные растения. Они имеют одну–две почки у корневой шейки, хорошо защищенные от зимних холодов и приспособленные к наиболее суровым зимним условиям местности. Эти растения не теряют отмерзающих побегов и ростков, зато весной развиваются медленнее протогемикриптофитов.
Криптофиты (Cr) – почки возобновления на подземных органах (клубнях, корневищах), скрыты в почве (геофиты) или под водой (гидрофиты и гедатофиты). Лучше всего защищены от иссушения. Многолетние травы с отмирающими надземными частями (ландыш, адонис амурский, хохлатки расставленная и сомнительная, василисники, ветровочники, луки, тюльпаны, лилии, калужница, такла, белокрыльник)
Эта жизненная форма подразделена на три подтипа:
- Геофиты. К этому подтипу отнесены растения, у которых почки и окончания побегов, приспособленные к перенесению неблагоприятного сезона, развиваются на подземных побегах на некоторой глубине. Особенно типичны они для степей, хотя встречаются и в других зонах, и там, где неблагоприятный период вызван засухой, и там, где он вызван морозами. Обычно растения этого подтипа имеют запасы питательных веществ.
Среди геофитов различают следующие группы растений:
Корневищные геофиты, имеющие более или менее удлиненные, обычно горизонтальные корневища (виды купены – Polygonatum, спаржи – Asparagus, вороньего глаза – Paris, ситника – Juncus, некоторые осоки – Сагех, злаки, например пырей ползучий – Agropyroп repens и тростник – Phragmites communis, ветреницы – Аnеmоnе, и др.).
Клубневые геофиты, имеющие клубни, служащие как для запасания питательных веществ, так и для перенесения неблагоприятных условий. Клубни могут иметь стеблевое происхождение, например, у цикламена – Cyclameп, хохлатки полой – Corydalis cava, очитка наибольшего – Sedum maximum, картофеля – Solаnиm tuberosum, земляной груши – Helianthus tuberosus и др.), корневое (например, у таволги шестилепестной – Filiреndulа hexapetala, пеона тонколистного – Paeonia tenuifolia, некоторых видов лютика – Ranunculus) и двойственное (в этом случае в их состав кроме корня, образующего большую часть клубня, входит и почка; примеры – многие орхидные, чистяк весенний – Ficaria verna и др.).
Клубнелуковичные геофиты (Раункиер этой группы геофитов не выделял), имеющие клубнелуковицу (клубнелуковица – видоизменение клубня, который несет в верхней части зачатки ассимилирующих листьев и окутан пленчатыми и волокнистыми основаниями высохших листьев); примеры: шафран (Сгоcus), шпажник (О ladiolus).
Луковичные геофиты. Запасают питательные вещества в чешуйчатых листьях, образующих луковицу. Луковица несет и почки, предназначенные для перенесения неблагоприятного периода. К этой группе геофитов относят лук (Аllium), птицемлечник (Ornithogalum), гусиный лук (Gagea), глоксинию (Gloxinia), тюльпан (Тuliра), нарцисс (Narcissus) и др.
Корневые геофиты. Переносят неблагоприятный период при помощи почек, располагающихся на сохраняющихся частях корней, остальные же органы растения, в том числе и верхние части корней, отмирают в начале неблагоприятного периода. К ним относятся вьюнок полевой (Coпvolvulus arveпsis), бодяки щетинистый и полевой (Cirsium setosum, С. arveпse) и др.
Переходную группу от гемикриптофитов к корневым геофитам представляют такие растения, как льнянка обыкновенная (ипаria vulgaris), щавелек (Rumex acetosella), осот желтый полевой.
(Soпchus arveпsis) и др., которые, будучи гемикриптофитами, в неблагоприятные годы, когда отмирают не только надземные органы, но и верхние части корней, сохраняются за счет почек на корнях, расположенных на некоторой глубине в почве.
- Гелофиты. К этому подтипу отнесены виды, которые растут в почве, насыщенной водой, или в воде, над которой поднимаются их листоносные и цнетоносные побеги. К их числу относятся аир (Acorus calamus), ежеголовка (Spargaпium), рогоз (Typha), камыш (Scirpus), частуха (Alisma), стрелолист (SagUtaria) и др.
- Гидрофиты. В этот подтип входят растения, живущие в воде и переносящие неблагоприятный период при помощи почек на корневищах или почек, свободно лежащих на дне водоема. Листья этих растений погруженные или плавающие; над поверхностью воды поднимаются (и то не у всех видов) только цветки или соцветия.
Среди них различают:
а) растения с корневищами на дне водоема, на которых находятся зимние почки (кувшинка – Nyтphaea, кубышка – Nuphar, многие рдесты – Potaтogetoп, элодея канадская – Elodea caпadeпsis и др.).
б) растения, на зиму полностью отмирающие, за исключением зимующих почек или коротких побегов, падающих на дно водоема
(пузырчатка – Utricularia, рдесты – Potaтogetoп, телорез Stratiotes aloides и др.).
Терофиты (Th) – возобновление только семянами. Неблагоприятный период года переживают на стадии семян. Все терофиты – однолетние растения (маки. марьянник розовый).
Ими сравнительно богаты степи, полупустыни и пустыни. К этой группе, кроме обычных однолетников, относятся и зимующие однолетники, которые, начав развитие осенью, зимуют в вегетативном состоянии и будущей весной или летом заканчивают свой цикл развития, давая семена.
Из перечисленных пяти основных жизненных форм наиболее примитивной, родоначальной формой следует считать ту, которая господствовала на Земле в период, когда климатические пояса и зоны еще не были выражены. В это время климат Земли, видимо, очень мало отличался от климата современных влажных тропических лесов и, следовательно, первичной формой следует считать фанерофитов с незащищенными почками, ныне господствующих в этих лесах. Постепенно условия существования растений на земном шаре стали дифференцироваться по количеству влаги, по продолжительности сухого и влажного периодов, по температурному режиму.
В разных географических условиях фанерофиты вырабатывали неодинаковые приспособления к неблагоприятному периоду. Многие из них выработали приспособления, защищающие листья в почках. В более суровых условиях ряд фанерофитов утратил вечнозеленость, и листья растений этой группы стали опадать на сухой или на холодный период года. В пределах группы фанерофитов возникли нанофанерофиты, затем низкорослые хамефиты и, наконец, гемикриптофиты, сохраняющие на неблагоприятный период лишь нижние части побегов, защищенные почвой или опавшими листьями. Возникновению групп геофитов и терофитов способствовали условия жарких сухих стран, где растения за короткий период с благоприятными условиями успевают принести плоды. Терофиты распространились здесь в связи со слабой сомкнутостью растительного покрова.
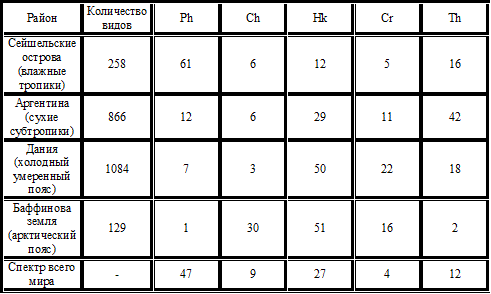
Процентное соотношение видов, обладающих разными жизненными формами на исследуемой территории, называется биоморфологическим спектром. Он служит своего рода индикатором условий окружающей среды и климата. В разных частях и странах земного шара спектры сильно различаются (таблица 3).
Как следует из таблицы, во влажных тропиках большинство видов относится к фанерофитам (и эпифитам), в сухом и жарком климате субтропиков – к терофитам, в холодном климате умеренного пояса – к гемикриптофитам, а в Арктике – к хамефитам. Но, несмотря на то, что отчетливо выражено соответствие типов жизненных форм географическим районам, они слишком обширны и неоднородны даже с учетом выделения в них подразделов. Таблица 3 – Биологический спектр
Особую группу среди жизненных форм составляют растения, которые присутствуют одновременно во многих ярусах. Это эпифиты – растут на других растениях (лишайники, мхи, хмель, плющ). Форма лианы – со слабым стволом, обвивающим другие растения, может быть как у древесных, так и у травянистых видов. К ним относятся виноград, хмель, луговая повилика, тропические лианы. Обвивая стволы и стебли прямостоячих видов, лиановидные растения выносят свои листья и цветки к свету (рис. 45).
Рисунок 45 – Растения–лианы – хмель
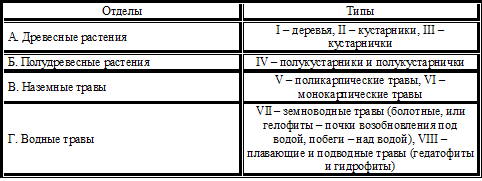
Сложный порядок соподчиненных классификационных признаков в середине XX века был предложен И.Г. Серебряковым (1964). Все многобразие растений сведено в 4 отдела и 8 типов жизненных форм (таблица 4), а каждый тип, в свою очередь подразделяется на формы.
Таблица 4 – Жизненные формы по И.Г. Серебрякову (1964)
На основе эколого–морфологического принципа была разработана классификация жизненных форм покрытосемянных растений (рис. 46). Именно она признана наиболее удачной для описания растительных сообществ.
Деревья с прямостоячими стволами – жизненная форма распространена очень широко и является показателем оптимальных условий местообитания. С ухудшением условий сменяется другими формами. Стланцы (кедровый стланик, ольховый стланик, сосна горная) растут в районах, мало благоприятных для произрастания деревьев – на Крайнем Севере, в предгольцовом горном поясе, на побережье Охотского моря, т.е. в районах с прохладным сырым летом, длинной зимой, обильными снегопадами, сильными ветрами. Деревья бывают первой (выше 25 м), второй (высота 10-25 м), и третьей (до 10 м) величины.
Кустарники – имеют множество одревесневших побегов, по высоте делятся на высокие (выше 2,5 м), средние (1-1,25 м), низкие (до 1,0 м).
Кустарнички – это те же кустарники, но никогда не превышающие 0,5 м. Обычно высота их составляет 10-30 см. Кустарнички доминируют в тундровых экосистемах, в напочвенном покрове северотаежных лесов, в горно–альпийском поясе (брусника, черника, голубика, подбел, низкие рододендроны, багульники и др.).
Полукустарники и полукустарнички – промежуточная форма между древесными растениями и травами. По облику сходны с травами, но в особо благоприятные годы стебли у них не отмирают, а продолжают на следующий год расти. Для них характерно сильное обмерзание побеговых систем. Типичный полукустарник – полынь Гмелина.

Рисунок 46 – Примеры жизненных форм покрытосеменных растений по классификации И.Г. Серебрякова (по И.Н. Пономаревой, 1975)
Травы – самый распространенный тип жизненной формы. Поликарпические травы – их большинство, плодоносят многократно. Они сильно различаются по строению корневых систем, отражающих их приспособленность к разным почвенным условиям. По этому признаку выделяют стержнекорневые (мятлики), длинностержневые (люцерна, шалфей), короткостержневые (сон–трава, крестовник), кистекорневые (калужница болотная, лютики), короткокорневищные (купена, ветровочник), длиннокорневищные (аспарагус), дерновинные (плотнокустовые, рыхлокустовые), столонообразующие (майник двулистный, земляника, сердечник белоцветковый, пырей), ползучие (вероника лекарственная, клевер белый, василисники), клубнеобразующие (аризема амурская, василисник клубненосный, звездчаточка лесная), луковичные (луки, хохлатки расставленная, сомнительная, Буша).
Монокарпические травы плодоносят всего один раз в жизни. Широко распространены в засушливых районах северного полушария. Все однолетники и двулетники (капуста, редисы, свекла, укроп, тмин, из дикой флоры: хохлатка бледная, пастушья сумка, чистотел, череда, борщевик, дудники даурский и амурский, донтестемон зубчатый и др.), есть и многолетники. Однолетники делятся на длительно вегетирующие (марьянник розовый, мак альпийский, офелия, пастушья сумка, кошачья лапка, патриния скабиозолистная) и эфемеры (вероника весенняя, горечавка Цолингера), лиановидные (горец вьюнковый), полупаразитные (очанка) и паразитные (вертляница одноцветковая, повилика, пучкоцвет трубкоцветковый).
Подушковидные формы – среди многолетних травянистых и древесных растений в особенно суровых условиях произрастания. Сугубо экологическая адаптация к холоду и засухе (камнеломки, вересковые, ива чукотская, дриады, рододендрон мелкоцветковый и др.)
Среди трав принято выделять группы, которые различаются физиономически, поскольку они играют разную экологическую роль.
Крупнотравье и папоротники – растут в наиболее влажных, но с плодородными, хорошо дренированными почвами, в большинстве своем теневыносливы. Высота от 1,0 до 2,0 м и более. Для растений типичны крупные, сложные листья, у цветковых видов развиты мощные стебли. Наиболее теплолюбивы в сравнении с другими видами. Характерны для широколиственных и хвойно–широколиственных, особенно пойменных, лесов. Особенно обильны виды этой группы в приокеанической полосе: В Южном Приморье, на юге Камчатке, на островах Японского моря.
Лесные виды: дудники, акониты, пионы, крапива, лабазник, клопогоны, стеблелист, недотрога обыкновенная, волжанка азиатская, чемерицы, чина Давида из папоротников – щитовники, корноптерис, кочедыжники, осмунд или чистоуст, и др. Луговые виды: купальница китайская, водосбор, ирисы, василисник вонючий, бузульник Фишера, соссюреи, подмаренник настоящий и др.
Разнотравье экологически сходно с первой группой, но более холодоустойчиво с продвижением на север сменяет первую группу. По строению растений не отличается от крупнотравьем, только уступает в размерах – не превышает 0,5 м, обычно 30-40 см.
Типичное разнотравье: подлесник красноцветковый, купена обертковая, звездчатка Бунге, марьянник розовый, чина низкая, молочай Комарова, вика однопарная, глухая крапива, диспорум зеленеющий, смилацина китайская.
Низкотравье – самые мелкие растения – до 20 см высотой, чаще 10-15 см. Самые теневыносливые, характеризуются высокой вегетативной подвижностью. Даже в глубокой тени могут образовывать группировки: тригонотис корейский, кислица обыкновенная, звездчаточка лесная, майники, седмичник, хохлатки, ветреницы, шлемник уссурийский. Многие виды разнотравья сугубо борельные виды и растут в северных лесах, но и на севере предпочитают более теплые экотопы с влажными плодородными почвами: седмичник европейский, майник двулистный, ветреница худосочная, грушанки мясокрасная и малая.
Злаки и осоки резко отличаются от типичных трав простыми линейными, преимущественно узкими листьями. В условиях достаточной освещенности сильно задерняют почвы. Некоторые виды выступают доминантами напочвенного покрова в разреженных лесах по всему Дальнему Востоку: вейник Лангсдорфа, мискантус краснеющий, осоки бледная, возвратившаяся, Ван–Хьюрка, и др. В данной группе имеются как ярко выраженные ксерофиты (осока низкая, овсяница овечья, мятлики), так и мезогигрофиты (камыш, осока мочажинная, тонконог, осока кривоносая).
Жизненные формы животных. Классификации жизненных форм животных, как и растений, весьма разнообразны и зависят от принципов, которые кладут в их основу. На морфологии млекопитающих, по–видимому, в наибольшей степени сказывается характер их передвижения в разных средах. А. Н. Формозов выделил на этом основании среди зверей следующие адаптивные типы: - наземные формы;
- подземные (землерои);
- древесные;
- воздушные и
- водные.
Между типами есть переходы.
В пределах каждой группы особенности поступательного движения и образа жизни формируют более специфические приспособительные формы. Так, наземные млекопитающие передвигаются в основном посредством ходьбы, бега и прыжков, что проявляется в их внешнем облике. Например, прыгающие животные: кенгуру, тушканчики, прыгунчики, кенгуровые крысы – отличаются компактным телом с удлиненными задними конечностями и значительно укороченными передними. Очень сильно развиты мышцы – разгибатели спины, увеличивающие мощность толчка. Длинный хвост играет роль балансира и руля, позволяющего резко изменять направление движения. Он служит также дополнительной точкой опоры. Сходный тип передвижения и внешний облик имели и некоторые вымершие динозавры мезозоя. Естественно, что, кроме общих адаптивных признаков, каждый вид отличается деталями внешнего строения в зависимости от экологической специфики.
Жизненные формы отчетливо выделяются в пределах любой крупной таксономической группы животных, характеризующейся экологическим разнообразием видов (рис. 47, 48).
Во внешнем облике птиц в наибольшей мере проявляется приуроченность их к определенным типам местообитаний и характер передвижения при добывании пищи. В связи с этим различают птиц: - древесной растительности;
- открытых пространств суши;
- болот и отмелей;
- водных пространств.
В каждой из указанных групп выделяют специфические формы: а) добывающие пищу посредством лазания (многие голуби, попугаи, кукушки, дятловые, воробьиные); б) добывающие пищу в полете (в лесах – совы, козодои и др., на открытых пространствах – ржанковые, длиннокрылые, над водой – трубконосые и т. п.); в) кормящиеся при передвижении по земле (лесные – большинство куриных, казуары, киви и др., на открытых пространствах – страусы, нанду, журавлиные, на болотах и отмелях – голенастые, фламинго, некоторые воробьиные); г) добывающие пищу с помощью плавания и ныряния (пингвины, гагары, поганки, ряд трубконосых, большинство веслоногих и гусиных и др.).
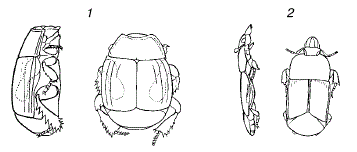
Рисунок 47 – Форма тела жуков-карапузиков из различных местообитаний (вид сбоку и сверху, по W. Kuhnelt, 1970): 1 – из навоза; 2 – из–под коры деревьев
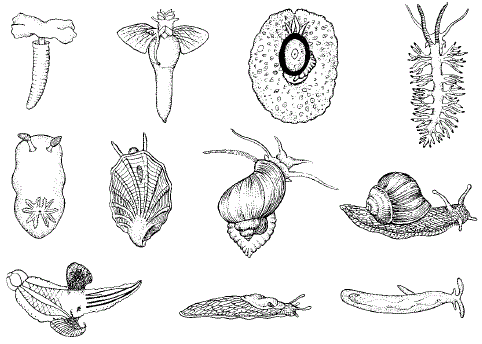
Рисунок 48 – Формы тела брюхоногих моллюсков, ведущих разный образ жизни
Жизненные формы таких мелких почвенных членистоногих, как коллемболы, выделяют на основе приуроченности их к определенным слоям почвенного профиля, так как в почве с глубиной резко меняется весь комплекс условий обитания: размеры полостей, освещенность, режим температуры и влажности и т. п.
Атмобионты – виды, населяющие верхние слои подстилки и способные подниматься на нижние части растений, характеризуются более крупными размерами, полностью развитыми глазами, относительно длинными придатками (антеннами, ногами и прыгательной вилкой), хорошо выраженной пигментацией. Эуэдафические виды – обитатели минеральных тонкопорозных слоев почвы – отличаются мелкими размерами, сильным укорочением придатков, редукцией вилки, полным отсутствием глаз и пигмента. Гемиэдафические формы характеризуются промежуточными чертами: частичной редукцией глаз, рассеянным пигментом, укороченной вилкой и др. (рис. 49).
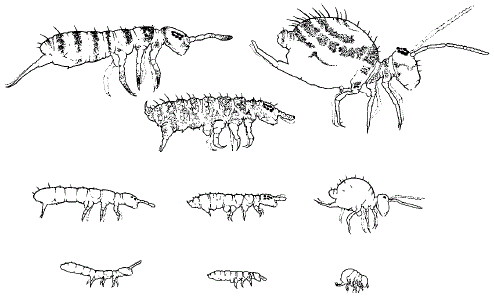
Рисунок 49 – Жизненные формы коллембол разных семейств (М. Б. Потапов)
Среди саранчовых по форме (конфигурация тела, структура головы, строение конечностей, летательного аппарата, тип покровительственной окраски) хорошо различаются тамнобионты – обитатели кустарников и деревьев; населяющие травянистый ярус хортобионты; жители надпочвенного слоя органических остатков герпетобионты; обитатели открытых участков грунта: эремобионты – на поверхности плотных глинистых почв, псаммобионты – на песках и петробионты – на каменистых участках с разреженной растительностью.
Сходные жизненные формы встречаются в сходных условиях жизни в разных зоогеографических областях и на разных материках (например, тушканчики Евразии, кенгуровые крысы Австралии, прыгунчики Африки). Жизненная форма наглядно свидетельствует об образе жизни вида.
Форма тела рыб, например, связана с тем, каким способом и с какой скоростью они передвигаются в воде – более плотной и вязкой среде, чем воздух. Торпедовидный тип тела имеют самые хорошие пловцы пелагиали (скумбрия, кефаль, лососи и др.). Виды со стреловидной, более вытянутой формой также приближаются к ним по скорости плавания в толще вод (сарган, щука). Сплющенное с боков тело – у рыб с меньшей скоростью, но такая форма позволяет хорошо маневрировать. Рыбы со змеевидным телом (угри, морские иглы и др.) обитают в зарослях. Морские коньки и тряпичники имитируют водоросли по форме. Лентовидное тело – сильно вытянутое и сплющенное с боков – у плохих пловцов (сельдяной король и т. п.). Рыбы, плавающие в придонных слоях, могут быть также шаровидными (кузовки), сплющенными дорзовентрально (скаты, морской черт). Движение рыб обеспечивается либо изгибанием всего тела, либо, при неподвижном теле, колебательными движениями разных плавников.
|