Тема 3.1. Структура популяций, видовая структура сообществ
3.1.1.Структурная организация популяций
Термин популяция (от лат. populus – народ, население) был предложен в 1903 году датским биологом В.Л. Иогансеном (1857– 1927).
По С.С. Шварцу, популяция – это элементарная группировка организмов определенного вида, свободно скрещивающихся и относительно изолированных от других особей того же вида, обладающая всеми необходимыми условиями для поддержания своей численности необозримо длительное время в постоянно изменяющихся условиях среды.
Согласно правилу объединения в популяции С. С. Четверикова, сформулировавшего его в 1903 году, индивиды любого вида живого всегда представлены не изолированными отдельностями, а их определенным образом организованными совокупностями. В связи с тем, что любая популяция обладает строго определенной генетической, фенотипической, половозрастной и другой структурой, она не может состоять из меньшего числа индивидов, чем необходимо для обеспечения стабильной реализации этой структуры и устойчивости популяции к факторам внешней среды. Именно в этом состоит принцип минимального размера популяций. Такой размер не есть константа для любых видов, он строго специфичен для каждой популяции. Выход за пределы минимума чреват для популяции гибелью: она уже не будет в состоянии самовозобновиться. Однако ее возможно восстановить искусственно, поскольку каждый индивид содержит в себе хотя и обедненное, но все же весьма значительное число генетических задатков вида, к которому он принадлежит.
Члены популяции характеризуются однотипными внешними признаками, сходными адаптациями и специфической реакцией на воздействие факторов среды, своеобразным типом динамики численности, демографической и территориальной структурой.
Популяция занимает двойственное положение в рядах биологических систем, принадлежащих разным уровням организации живой материи (рис. 82). С одной стороны популяция является одним из звеньев генетико–эволюционного ряда, отражающего филогенетические связи таксонов разного уровня, как результат эволюции форм жизни:
организм – популяция – вид – род – … – царство
В этом ряду популяция выступает, как форма существования вида, основная функция которого заключается в выживании и воспроизведении. Играя важную роль в микроэволюционном процессе, популяция является элементарной генетической единицей вида. Особи в популяции обладают характерными особенностями строения, физиологии и поведения, то есть гетерогенностью. Эти особенности вырабатываются под влиянием условий обитания и являются результатом микроэволюции, протекающей в конкретной популяции. Изменение популяций в процессе адаптации к изменяющимся факторам среды и закрепление этих изменений в генофонде обусловливает в итоге эволюцию вида. С другой стороны, в тех же конкретных условиях среды популяция вступает в трофические и иные связи с популяциями других видов, образуя с ними простые и сложные биогеоценозы. В этом случае она является функциональной субсистемой биогеоценоза и представляет одно из звеньев функционально–энергетического ряда:
организм – популяция – биогеоценоз – биосфера
«Двойственность» популяций проявляется и в биологической противоречивости их функций. Они сложены особями одного вида, а, следовательно, одинаковы по экологическим требованиям к условиям среды, и обладают одинаковыми механизмами адаптации. Но в себе самих популяции содержат высокую вероятность острой внутривидовой конкуренции и возможность отсутствия устойчивых контактов и взаимосвязей между особями.

Рисунок 82 – Положение популяции в структуре биологических систем биосферы (по И.А. Шилову, 1988)
Острая конкуренция имеет место при перенаселении, ведущем к истощению жизнеобеспечивающих ресурсов: у животных – пищи, у растений – влаги, плодородия и (или) света. При слишком малой численности особей популяция утрачивает свойства системы, устойчивость ее снижается. Разрешение данного противоречия является главным условием сохранения целостности системы. Оно заключается в необходимости поддержания оптимальной численности и оптимального соотношения между внутрипопуляционными процессами дифференциации и интеграции.
Степень обособленности популяций. Если члены вида постоянно перемещаются и перемешиваются на обширных пространствах, такой вид характеризуется небольшим числом крупных популяций. Большими миграционными способностями отличаются, например, северные олени и песцы. Результаты мечения показывают, что песцы перемещаются за сезон на сотни, а иногда более чем на тысячу километров от мест размножения. Северные олени совершают регулярные сезонные кочевки также в масштабах сотен километров. Границы между популяциями таких видов проходят обычно по крупным географическим преградам: широким рекам, проливам, горным хребтам и т. п. В некоторых случаях подвижный вид при относительно небольшом ареале может быть представлен одной-единственной популяцией, например кавказский тур, стада которого постоянно кочуют по двум основным хребтам этого горного массива.
При слабо развитых способностях к перемещению в составе вида формируется множество мелких популяций, отражающих мозаичность ландшафта. У растений и малоподвижных животных число популяций находится в прямой зависимости от степени разнородности среды. Например, в горных районах территориальная дифференцировка таких видов всегда более сложна, чем на ровных открытых пространствах. Примером вида, у которого множественность популяций определяется не столько дифференцировкой среды, сколько особенностями поведения, является бурый медведь. Медведи отличаются большой привязанностью к местам своего обитания, поэтому в пределах обширного ареала представлены множеством относительно мелких группировок, отличающихся друг от друга по ряду свойств.
Степень обособленности соседних популяций вида очень различна. В некоторых случаях они резко разделены территорией, непригодной для обитания, и четко локализованы в пространстве, например популяции окуня и линя в изолированных друг от друга озерах или популяции пластинчатозубой крысы, белоусой славки, индийской камышевки и других видов в оазисах и долинах рек среди пустынь.
Противоположный вариант – сплошное заселение видом обширных территорий. Такой характер распространения свойствен, например, малым сусликам в сухих степях и полупустынях. В этих ландшафтах плотность их населения повсеместно высока. Отдельные непригодные для жизни участки легко преодолеваются при расселении молодняка, а в благоприятные годы на них возникают временные поселения. Здесь вычленить границы между популяциями можно лишь условно, между областями с разной плотностью населения.
Другой пример сплошного распространения вида – семиточечная божья коровка. Эти жуки встречаются в самых различных биотопах и разных природных зонах. Вид характеризуется, кроме того, предзимними миграциями. Границы между популяциями в таких случаях почти не выражены. Однако, поскольку совместно обитающие особи контактируют между собой чаще, чем с представителями других частей ареала, население удаленных друг от друга мест можно считать разными популяциями.
В пределах одного и того же вида могут быть популяции как с хорошо различимыми, так и со смазанными границами. У вредителя зерновых культур на юге нашей страны – клопа–черепашки (Eurygaster integriceps) – через 2,5–3 месяца после выхода из яиц и активного питания на полях начинается отлет в леса предгорий, за десятки, а порой и сотни километров, где клопы залегают до следующей весны под слой древесного опада. Дальность перелетов зависит от степени упитанности особей, и в результате на одних и тех же зимовках смешиваются клопы из разных мест. Направление весеннего разлета сильно зависит от направления ветра, и клопы не обязательно возвращаются в места отрождения. Кроме того, часть черепашек не улетает далеко, а зимует в ближайших лесополосах. Таким образом, на обширных территориях происходит постоянное перемещение и перемешивание особей, которое не позволяет строго разграничивать отдельные популяции. Между тем в определенных частях ареала, как горных, так и равнинных, черепашка ведет относительно оседлый образ жизни, совершая лишь короткие перелеты к ближайшим местам зимовок. В таких районах возникают обособленные популяции, представляющие собой отдельные локальные очаги размножения вредителя. Таким образом, в пределах вида популяции могут быть представлены группами разного объема.
Между популяциями осуществляется обмен отдельными особями, который может быть либо достаточно регулярным, либо эпизодическим. При сезонных кочевках ворон, например, часть молодых птиц ежегодно остается на местах зимовок, образуя пары с представителями оседлого населения. Связь между населением отдельных видов рыб в озерах осуществляется значительно реже, например в годы с особо сильными паводками, когда обособленные водоемы соединяются в единую водную систему.
Взаимоотношения видов не сводятся только к взаимодействию особей, хотя и осуществляются через индивидуальные контакты. Отношения между хищником и его жертвой кратковременны. Связь между их популяциями длительна и является одним из условий функционирования сообществ. Связи между отдельными особями приводят к одним результатам, связи между популяциями – к качественно другим. Например, длительное воздействие конкретного паразита может иметь следствием изменение физиологического состояния, плодовитости, долговечности хозяина и т. п. Взаимоотношения популяций этих же видов приводят к изменениям их основных групповых характеристик: численности, возрастного состава, смертности, темпов роста популяций.
Связи между популяциями поддерживают вид как единое целое. Слишком длительная и полная изоляция популяций приводит обычно к образованию новых видов.
Различия между отдельными популяциями выражены в разной степени. Они могут затрагивать не только их групповые характеристики, но и качественные особенности физиологии, морфологии и поведения отдельных особей. Эти различия создаются в основном под влиянием естественного отбора, приспосабливающего каждую популяцию к конкретным условиям ее существования.
По способности к самовоспроизведению различают постоянные и временные популяции. Постоянные популяции могут быть независимыми, не нуждающимися в притоке особей извне для поддержания своей численности, и полузависимыми, когда такой приток существенно повышает численность популяции, но она может сохраняться и без него. Временные популяции могут также быть различного типа. Общие их особенности в том, что собственный приплод не покрывает смертность и длительность их существования зависит от мигрантов. По размерам различают карликовые, обычные локальные и суперпопуляции. Последние занимают сплошь обширные территории и состоят из большого числа особей. Единство таких популяций в основном топографическое, так как взаимные контакты особей ослаблены расстоянием. В их пределах вычленяют субпопуляции разных масштабов.
Широкое распространение в экологии получила классификация популяций по Н.П. Наумову в зависимости от размеров занимаемой ими территории. В этой классификации различают элементарную, экологическую и географическую популяции (рис. 83).
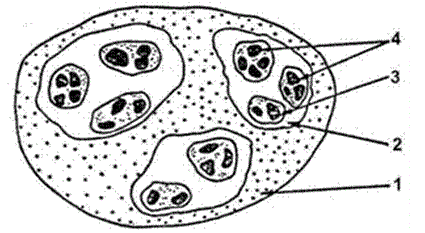
Рисунок 83 – Пространственная иерархия популяций (по Н.П. Наумову, 1963):
- – ареал вида, популяции;
- – элементарная;
- – экологическая;
- – географическая
Элементарная (локальная) популяция – это совокупность особей одного вида, занимающих небольшой участок однородной площади. Между ними постоянно идет обмен генетической информацией. К локальным популяциям относятся, например: микрогруппировки ландыша Кейске в березняке, растущие у оснований деревьев и на открытых местах; одна из нескольких стай рыб одного вида, обитающего в озере; куртины деревьев одного вида (дуба монгольского, лиственницы, и др.), разобщенные лугами, куртинами других деревьев или кустарников, или болотцами. Количество элементарных популяций, на которые распадается вид, зависит от разнородности условий в биогеоценозе: чем они однообразнее, тем меньше элементарных популяций и наоборот.
Экологическая популяция – совокупность элементарных популяций, внутривидовые группировки, приуроченные к конкретным биоценозам. Растения одного вида в ценозе называются ценопопуляцией. Обмен генетической информацией между ними происходит достаточно часто. Примерами являются рыбы одного вида во всех стаях общего водоема; популяции белок в сосновых, елово-пихтовых и широколиственных лесах одного района.
Географическая популяция – совокупность экологических популяций, заселивших географически сходные районы. Географические популяции существуют автономно, ареалы их относительно изолированы, обмен генами происходит редко, так например, у животных и птиц – во время миграций, у растений – при разносе пыльцы, семян и плодов. На этом уровне происходит формирование географических рас, разновидностей, выделяются подвиды. Известны географические расы лиственницы даурской (Larix dahurica): западная (к западу от р. Лены) (L. dahurica ssp. dahurica) и восточная (к востоку от р. Лены, выделяемая в L. dahurica ssp. cajanderi), северная и южная расы лиственницы курильской. Аналогично выделение М.А. Шембергом (1986) у березы каменной двух подвидов: березы Эрмана (Betula ermanii) и шерстистой (B. lanata). Зоологи выделяет тундровую и степную популяции у узкочерепной полевки (Microtis gregalis): тундровые более крупные, значительно раньше начинают размножаться, обладают более высокой плодовитостью и больше накапливают жира. Вид «белка обыкновенная» насчитывает около 20 географических популяций, или подвидов. Зайцы–беляки из разных частей ареала различаются характером окраски, размерами, строением пищеварительной системы. Например, длина слепой кишки у зайцев полуострова Ямал в 2 раза больше, чем у представителей из лесостепного Урала. Это связано с характером питания, разной долей грубых кормов в составе рациона. Средняя величина выводка в центрально–якутской популяции составляет около 7 зайчат на самку, которая дает приплод один раз в год. В карельской части ареала зайчихи приносят зайчат дважды за сезон, но средняя величина выводка около 4 экземпляров. В Белоруссии самки беляков могут давать до четырех приплодов в год. Заячье население разных географических территорий испытывает неодинаковое давление со стороны хищников и паразитов. Например, в Верхоянье до 70 % зайцев бывает поражено цестодами, вызывающими заболевание мозга. Архангельская популяция больше других страдает от печеночного сосальщика. Различия в характере размножения и масштабах гибели от тех или иных причин определяют различия в общей численности, плотности, темпах роста и структуре популяций разных регионов. Популяции различаются между собой тем сильнее, чем более несходны условия их обитания и чем слабее между ними обмен особями.
Территориальные границы популяций разных видов не совпадают. Наряду с видами, у которых более или менее отчетливо вычленяются локальные популяции (мышевидные грызуны и другие мелкие млекопитающие, некоторые немигрирующие насекомые, моллюски и т. п.), существуют виды с более широкими популяционными границами. Например, территория, занимаемая одной популяцией таких крупных подвижных животных, как лоси, включает множество конкретных участков с разнообразной растительностью: леса с господством различных пород, поля, луга, овраги, поймы рек и т. п. Не ограничены местными пределами популяции многих птиц, крупных млекопитающих, а также ряда мелких организмов с эффективными способами расселения.
Многообразие природных популяций выражается также в многообразии типов их внутренней структуры.
Структура популяции – это состав особей и их распределение в пространстве. Сложность структуры зависит от соотношения возрастных групп, полов, наличия различного рода группировок, территориальных взаимоотношений. Чем сложнее структура популяции, тем выше ее приспособительные возможности.
Пространственная структура популяции – характер распределения особей в пределах ареала, зависящий от особенностей окружающей среды. В природе изредка встречается почти равномерное упорядоченное распределение особей на занимаемой территории, например в уплотненных популяциях сидячих морских полихет, в чистых зарослях некоторых растений. Однако чаще всего члены популяции распределяются в пространстве неравномерно, что обусловлено двумя причинами: во-первых, неоднородностью занимаемого пространства, а во-вторых, некоторыми особенностями биологии видов, которые способствуют возникновению скоплений индивидуумов. У растений такая агрегированность возникает, например, при вегетативном размножении, при слабом распространении семян и прорастании их вблизи материнской особи; у животных – при групповом образе жизни семьями, стадами, колониями, при концентрациях для размножения, перезимовки и т. п.
Неравномерное размещение членов популяции может проявляться в двух крайних вариантах со всевозможными переходами между ними: - в резко выраженной мозаичности с незанятым пространством между отдельными скоплениями особей и
- в распределении случайного, диффузного типа.
Примером первого могут служить гнездовья грачей, поселяющихся колониями в рощах или парках, к которым примыкают благоприятные кормовые угодья. Диффузное распределение встречается в природе, если члены популяции относительно независимы друг от друга и обитают в однородной для них среде. Таково, например, размещение мучных хрущаков Tribolium confusum в муке, личинок-поденок в воде ручья, норок пауков–каракуртов на лугах и т. п.
В каждом конкретном случае тип распределения в занимаемом пространстве оказывается приспособительным, т. е. позволяет оптимально использовать имеющиеся ресурсы. Способы, которыми достигается рациональный характер размещения, определяются системой взаимоотношений между членами популяции.
Растения в ценопопуляции чаще всего распределены крайне неравномерно, образуя более или менее изолированные группы, скопления, так называемые микроценопопуляции, субпопуляции или ценопопуляционные локусы. Эти скопления отличаются друг от друга числом особей, плотностью, возрастной структурой, протяженностью. Часто более плотный центр скопления окружен особями, расположенными менее плотно.
Пространственная неоднородность ценопопуляции связана с характером развития скоплений во времени. У люцерны серповидной, например, семена обычно опадают в непосредственной близости от материнского растения, поэтому около обильно плодоносящих средневозрастных генеративных растений образуются скопления молодых (проростков, ювенильных и имматурных). Эти скопления характеризуются большой плотностью. По мере того как особи переходят в следующие возрастные состояния, скопления меняют возрастную структуру, изреживаются. Одновременно происходит приживление новых зачатков в пределах скопления, оно уплотняется, усложняется его структура, расширяется занятая им территория. Если приживление зачатков происходит за пределами скопления, то возникает новое. Скопления могут частично сливаться друг с другом, т. е. достигать более высокого уровня агрегированности.
Таким образом, жизнь ценопопуляции протекает в форме несинхронных возрастных изменений различных локусов, при этом меняется и ее пространственная структура, ибо изменяются конфигурация, протяженность локусов и их местоположение в фитоценозе.
Определить характер размещения растений можно с помощью простейшей математической обработки данных учета. Участок или пробную площадь разбивают на учетные площадки одинакового размера – не менее 25, или же проводят учеты растений на расположенных примерно на одном и том же расстоянии учетных площадках одинакового размера. Совокупность площадок представляет собой выборку. Обозначив среднее число особей вида на площадках в выборке буквой m, количество площадок (учетов) в выборке – n, фактическое число особей вида на каждой площадке – x, можно определить дисперсию, или меру рассеяния S2 (отклонение значения x от m):
S2 = S(m-x)*2 /(n-1 )
При случайном распределении S2 = m (при условии достаточного размера выборки). При равномерном распределении S2 = 0, а число особей на каждой площадке должно быть равным среднему. При групповом распределении всегда S2 > m, и чем больше разница между отклонением и средним числом, тем сильнее выражено групповое размещение особей.
У животных благодаря их подвижности способы упорядочивания территориальных отношений более разнообразны по сравнению с растениями. Даже у сидячих форм имеется ряд приспособлений к рациональному размещению в пространстве. У асцидий и мшанок растущий край колонии, наталкиваясь на колонию другого вида, растет поверх нее и в конечном счете подавляет ее. Но если встречаются колонии одного вида, каждая из них тормозит рост соседней и они начинают распространяться в другом направлении. При полном окружении колониями своего вида вегетативное размножение прекращается, но усиливается образование половых продуктов и подвижных личинок.
Личинки устриц, морских желудей, сидячих полихет, руководствуясь химическими стимулами, оседают обычно в местах, где уже имеются особи того же вида. Для осевшей личинки перед окончательным прикреплением характерно специальное поисковое поведение, позволяющее ей занять определенное место в скоплении. По наблюдениям, циприсовидные личинки морского желудя Balanus balanoides обычно прикрепляются на расстоянии не менее 2,5 мм от молодых и не менее 2 мм от старых особей своего вида. В то же время они свободно оседают в близком соседстве и даже на поверхности представителей других видов.
У высших животных внутрипопуляционное распределение регулируется системой инстинктов. Им свойственно особое территориальное поведение – реакция на местонахождение других членов популяции. Инстинкты, поддерживающие размещение по территории отдельных особей или групп в популяциях, существуют у птиц, млекопитающих, пресмыкающихся, ряда рыб, в меньшей степени – у амфибий. Они выражены также у многих видов беспозвоночных со сложной нервной системой – ряда насекомых, пауков, крабов, осьминогов и др.
По типу использования пространства все подвижные животные подразделяются на оседлых и кочевых. При оседлом существовании животное в течение всей или большей части жизни использует довольно ограниченный участок среды. Такие животные отличаются инстинктами привязанности к своему участку, а в случае вынужденного переселения – стремлением вернуться на хорошо знакомую территорию. Это «чувство дома» получило в экологии название «хоминг» (от англ. home– дом). Многие виды возвращаются к месту размножения даже после длительных и дальних миграций. Известно, например, что одна и та же пара скворцов из года в год может занимать «свой» скворечник. Хоминг голубей использовали даже в практических целях – для переноса почты.
Оседлый образ жизни имеет значительные биологические преимущества. На хорошо знакомой территории животное свободно ориентируется, тратит меньше времени на поиски корма, кратчайшим путем спасается в известные ему укрытия. Кроме того, многие оседлые виды создают систему запасов, прокладывают тропинки, строят дополнительные гнезда, норы, что помогает им выжить. Например, у белок есть основное гнездо, где выводится молодняк, и несколько дополнительных, в которые зверьки скрываются от непогоды или врагов. Белки создают также серии кладовых, запасая на зиму орехи, семена, грибы. Длительное использование определенной территории помогает более полному ее освоению.
На чужом участке поведение животного меняется. Наблюдения за сусликами показали, например, что такие зверьки отличаются суетливостью, неуверенностью передвижения, часто осматриваются, убежища находят лишь случайно и поэтому чаще гибнут, чем хозяева территории.
Однако оседлый образ жизни таит в себе угрозу быстрого истощения ресурсов, если плотность популяции окажется слишком высокой. У оседлых видов выработались такие приспособительные особенности поведения, которые обеспечивают разграничение мест обитания между отдельными особями, семьями или другими внутрипопуляционными группировками (рис.84). Общая площадь, занимаемая популяцией, оказывается поделена на отдельные индивидуальные или групповые участки, чем достигается упорядоченное использование запасов пищи, естественных укрытий, мест для размножения и т. п.

Рисунок 84 – Основные типы домовых участков у животных (по Н. П. Наумову, 1972):
А – одиночные разобщенные охраняемые;
Б – одиночные перекрывающиеся;
В – групповые разобщенные;
Г – групповые, перекрывающие друг друга (у колоний птиц на островах)
Территориальное поведение животных. Несмотря на территориальное обособление членов популяции, между ними поддерживается связь с помощью системы различных сигналов и непосредственных контактов на границах владений. Территориальное поведение животных включает два типа активности: направленную на обеспечение собственного существования (поиск пищи, исследование территории, рытье нор и т. п.) и - на установление отношений с соседними особями (охрана участков, сигнализация, мечение и др.).
Занимаемые участки не бывают больше тех пределов, при которых связь между соседними особями нарушается. Минимальные же размеры индивидуальной территории зависят от кормовых ресурсов среды. При такой системе размещения каждая оседлая особь не встречает конкуренции на своем участке, это повышает шансы на выживание и оставление потомства (рис. 85).
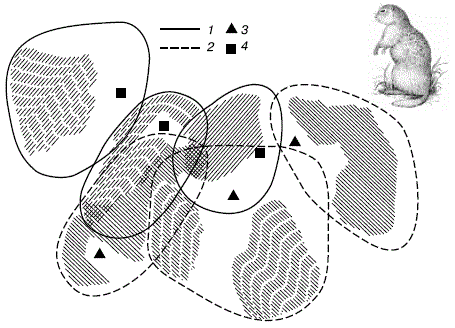
Рисунок 85 – Индивидуальные участки шести малых сусликов в Нижнем Поволжье (по А. Н. Солдатовой, 1955):
- границы индивидуальных участков у самок;
- то же у самцов;
- гнездовые норы самцов;
- выводковые норы самок.
Заштрихованы предпочитаемые каждым зверьком места кормежки
«Закрепление участка» достигается разными способами: - охраной границ занимаемого пространства и прямой агрессией по отношению к чужаку;
- особым ритуальным поведением, демонстрирующим угрозу; системой специальных сигналов и меток, свидетельствующих о занятости территории.
Прямая агрессия и нанесение конкуренту физических повреждений встречаются довольно редко. Перед нападением обычно демонстрируется поведение угрозы. Часто «схватка» представляет собой лишь систему ритуальных поз и заканчивается, как правило, изгнанием пришельца. В территориальных конфликтах в большинстве случаев побеждает хозяин участка, даже если ему противостоит более сильный соперник. В поведении животных на своей территории преобладает рефлекс активной обороны, на чужой – ориентировочный. Малый суслик немедленно бросается на другого, если тот забегает на его участок, и преследует нарушителя до границы своей территории. Если же, увлекшись погоней, он попадает на участок соседа, роли меняются, и хозяин активно изгоняет зверька, от которого только что спасался сам.
Такое же право первого осуществляется и в территориальных притязаниях у большинства птиц. Гнезда мухоловок–пеструшек в эксперименте передвигали в переносных дуплянках на гнездовые участки соседних пар. При этом возникали драки, в которых всегда побеждали хозяева участка. Если гнездо победившей пары оказывалось на территории соседей, исход драки решался в пользу владельцев участка.
Многие животные вообще редко вступают в конфликты, активно избегая чужие участки. Широко развита система маркировки, предупредительных сигналов о занятости территории. У птиц это в основном сигнализация звуками. Поющий самец зарянки, например, все время передвигается по границам своего участка, задерживаясь в тех местах, откуда грозит вторжение соседа. Млекопитающие часто оставляют запаховые метки: мочу, кал, выделения специальных желез. У сурков и многих других видов для мечения участка служат анальные железы. У соболей пахучий секрет выделяется на подошвах ног, у серн – позади рогов, у антилопы гарна – спереди от внутреннего края глаз. Серны и антилопы оставляют запаховые следы на кустарниках и нижних ветвях деревьев. Собачьи, кошачьи и лемуры метят территорию мочой. Лемуры для этой цели ладонями втирают капли мочи в подошвы ног.
Ряд крупных животных оставляют метки, воспринимаемые зрительно (рис. 86). Например, медведи и дикие коты оставляют глубокие царапины на деревьях.
Обычная реакция на территориальные метки – избегание – закреплена у животных наследственно. Биологическая выгода такого типа поведения очевидна. Если бы овладение территорией решалось только исходом физической борьбы, появление каждого более сильного пришельца грозило бы хозяину потерей участка и устранением от размножения. Большая часть энергии особей тратилась бы на постоянные стычки, что уменьшило бы шансы выведения потомства и увеличило смертность.

Рисунок 86 – Зубр, маркирующий дерево (по И. И. Барабаш-Никифорову, А. Н. Формозову, 1963)
Охрана участка может осуществляться не на всей его территории, а лишь вблизи биологического центра – норы, гнезда и т. п. В таких случаях владения отдельных особей или групп могут перекрываться, иногда широко, и часть пространства животные используют совместно. На нейтральной территории инстинкт агрессии угасает. Так, мухоловки-пеструшки, гнезда которых искусственно сближают на границе соседних участков, мирно живут рядом и выкармливают птенцов.
Частичное перекрывание индивидуальных территорий служит способом поддержания контактов между членами популяции. Соседние особи часто поддерживают устойчивую обоюдовыгодную систему связей: взаимное предупреждение об опасности, совместную защиту от врагов. Нормальное поведение животных включает активный поиск контактов с представителями своего вида, который часто усиливается в период падения численности.
Территориальное поведение животных особенно ярко выражено в период размножения. Оно тесно связано с их физиологическим состоянием и регулируется гормональным путем. Вне сезона размножения у многих видов распределение по индивидуальным участкам сменяется групповым образом жизни с иным типом поведения. Большинство воробьиных птиц после вылета птенцов объединяются в стаи, совершающие кочевки.
У ряда перелетных птиц территориальное поведение проявляется и на местах зимовок. Хищные и мелкие мышевидные грызуны обладают участками на протяжении всей жизни. Появившийся молодняк сначала остается на территории родителей, но затем выселяется, либо занимая освободившиеся участки, либо проникая в соседние популяции, либо формируя новые на незанятой территории.
В устойчивых семейных группах территориальные отношения между членами семьи могут быть достаточно сложными. Тигры, которым присуща полигамия, живут семьями, состоящими из самца, двух–трех самок и молодых разного возраста. Площадь семейной территории определяется плотностью основных жертв и при 14–15 особей копытных на 1000 га составляет около 50 тыс. га на семью. Территория, посещаемая самками, значительно меньше – около 10 тыс. га. Самец периодически обходит всю семейную территорию по постоянному маршруту, оставляя пахучие метки, экскременты и мочу на выступах скал, выворотах деревьев, нагребаемых кучах земли, снега и т. п. Участки молодых тигров частично перекрываются с территорией, посещаемой самками. Некоторые части семейных владений посещают лишь старые и молодые самцы. У каждого члена семьи есть излюбленные места для охоты. Через 3–5 лет молодые покидают семейную территорию родителей.
У оседлых видов все варианты общей пространственной структуры популяций можно свести к четырем основным типам: диффузному, мозаичному, пульсирующему и циклическому.
В популяциях диффузного типа животные распределены в пространстве дисперсно, не образуя обособленных поселений. Это возможно лишь при относительно равномерном размещении по территории кормов, мест, пригодных для размножения, и укрытий. Диффузный тип пространственной структуры популяций преобладает, например, у мелких млекопитающих сухих открытых пространств – большой песчанки и мохноногого тушканчика в монотонных песчаных ландшафтах, даурской пищухи в полынной и карагановой степи, малого суслика в различных растительных ассоциациях сухих степей и полупустынь.
Мозаичный тип размещения оседлых животных возникает в том случае, если пригодные для заселения биотопы распределены в пространстве резко неравномерно. Например, поселения малого суслика в волго–уральской полупустыне приурочены только к пятнам степной растительности, встречающимся среди сплошных песков. Обыкновенный хомяк в полупустыне обитает только в тростниковом поясе озер и на склонах озерных котловин.
Пульсирующий тип пространственной структуры характерен для популяций с резкими колебаниями численности. В годы депрессии популяция состоит из обособленных поселений, а в годы подъема численности занимает всю пригодную территорию, меняя мозаичный тип распределения на диффузный. В период резкого падения численности животные собираются на наиболее благоприятных участках, которые получили название «cтаций переживания». Например, полевки–экономки в лесостепи в засушливые годы заселяют только березово–осиновые колки и заболоченные берега озер. Во влажные годы возникает обширная сеть временных поселений практически во всех биотопах, включая посевы. Эти временные поселения сохраняются обычно лишь до следующей засухи.
Пульсирующий тип структуры популяций можно отметить и у многих видов панцирных клещей и коллембол, заселяющих лесную подстилку. В сухое время эти членистоногие концентрируются лишь возле корней деревьев, где режим влажности стабильнее, а во влажные периоды их популяции занимают весь покров растительных остатков на поверхности лесной почвы.
Циклический, или переложный, тип пространственной структуры популяций оседлых животных характеризуется закономерным попеременным использованием территории в течение года. Обский и копытный лемминги на острове Врангеля зимуют на сухих прибрежных возвышенностях. Летом зверьки переселяются на кочкарные и разнотравно–злаково–лишайниковые участки тундр, где и располагаются их летние норы. Лемминги отличаются высокой динамичностью пространственного распределения, отсутствием постоянных поселений, сменой мест зимнего и летнего обитания. Такой переложный характер использования территории в условиях тундр имеет большое приспособительное значение. Восстановление нарушенной растительности в тундрах происходит чрезвычайно медленно, длительное существование на одном месте за счет растительных кормов практически невозможно и грозило бы не только голодом зверькам, но и необратимыми изменениями растительного покрова. При переложном типе использования пространства сохраняется баланс между потреблением кормов и их ежегодным возобновлением, а тем самым и возможность существования дальнейших поколений.
Циклический тип освоения территории оседлыми животными, по существу, сходен с использованием ресурсов кочевыми популяциями.
Преимущества кочевого образа жизни заключаются в том, что животные не зависят от запасов корма на конкретной территории. Постоянные передвижения одиночных особей, однако, увеличивают вероятность гибели от хищников. Поэтому кочевой образ жизни практически несвойствен одиночным животным, кочуют группы: стада, стаи. Индивидуальные участки отдельных особей в таких группах не обособляются, но широко развита ориентация на соседей. В пасущихся стадах копытных между отдельными животными поддерживается определенное среднее расстояние, при котором они не мешают друг другу, обладая свободой передвижения. У птиц и рыб расположение в стае улучшает аэро– и гидродинамические условия полета и плавания: обеспечивается такое взаимодействие вихревых следов движения каждой особи, которое уменьшает общую величину сопротивления среды. У рыб, плывущих в середине стаи, частота движения хвостового плавника меньше, чем у тех, которые находятся впереди. С изменением скорости дистанция между членами движущейся стаи меняется.
Некоторые виды образуют широко кочующие группы, не привязанные к определенной территории. Таково поведение многих видов рыб во время нагульных миграций. Масштабы и длительность таких миграций зависят от обилия корма и численности стада. Чем крупнее стадо, тем длиннее его миграционный путь, так как быстрее потребляются встреченные корма. Норвежская сельдь Clupea harengus после нереста у берегов Норвегии кочует на тысячи километров на север и восток, держась в местах скопления планктона, который служит ей пищей.
В некоторых случаях, при резких нерегулярных изменениях запасов корма, возможны столь же нерегулярные перемещения популяции и временное «оседание» ее в тех местах, где запасы пищи еще не исчерпаны. Типичные примеры таких популяций без постоянных территорий можно обнаружить у клестов, всецело зависящих от неустойчивого урожая семян хвойных. У клестов постоянно меняются места гнездования и сроки размножения. В кормных лесах птицы выводят птенцов не только летом, но и зимой. Во внегнездовое время они широко кочуют в поисках лесов с богатым урожаем семян.
Перемещения многих других видов ограничены определенной территорией, которая может быть очень большой. Табуны зебр в Серенгети кочуют в сухой сезон на участке в 400–600 км2, во влажный общая площадь их перемещения занимает 300–400 км2. На этой территории у них есть определенные места отдыха, пастьбы и водопоев, которые они используют регулярно в определенные сроки. Периферические части таких территорий могут широко перекрываться с участками других стад. Регулярное перемещение по площади обычно происходит за сроки, достаточные для восстановления кормов на пастбищных участках. Сходным образом используют территорию стада других копытных, слонов, обезьян. Таким образом, при кочевом образе жизни животные совершают более или менее закономерные циклические перемещения по обширной территории с повторным использованием одних и тех же участков через определенные сроки.
У кочующих видов возможно также обособление оседлых популяций или временный переход к оседлому существованию. Северные олени на островах Северного Ледовитого океана зимуют там же, не делая попыток перехода на материк. Таймырские стада совершают обширные миграции более чем на 1000 км, но отдельные мелкие группы остаются на местах летнего нагула на северном Таймыре, как и на местах зимовок в лесотундре и северной части тайги.
Между разными способами использования территории нет абсолютных разграничений. Пространственная структура популяции очень динамична. Она подвержена сезонным и другим адаптивным перестройкам в соответствии с местом и временем. Однако масштабы возможных изменений и общий тип использования территории определяются биологическими особенностями вида, в первую очередь характером внутривидовых связей. Особенно большую роль в поддержании структуры популяций играет поведение особей по отношению друг к другу, система взаимных контактов.
Многочисленные конкретные формы проявления территориальных взаимоотношений животных можно расположить в постепенно усложняющийся ряд. Самая простая пространственная структура популяций – система неперекрывающихся охраняемых участков оседлых особей; более сложная – система перекрывающихся территорий, при которой усиливаются внутривидовые контакты. Наконец, групповое использование участков возможно только на основе строго упорядоченных отношений внутри группы.
Этологическая (поведенческая) структура популяций у животных обусловлена наличием внутрипопуляционных группировок, члены которых непосредственно взаимодействуют между собой. Такие группировки называют демы (семьи), соседства, субпопуляции, парцеллы, локусы.
Существует множество уровней агрегации особей: одиночные особи, семьи, колонии, стада, стаи.
Одиночный образ жизни при котором особи популяции независимы и обособлены друг от друга, характерен для многих видов, но лишь на определенных стадиях жизненного цикла. Полностью одиночное существование организмов в природе не встречается, так как при этом было бы невозможным осуществление их основной жизненной функции – размножения. Однако для некоторых видов характерны очень слабые контакты между совместно живущими особями. Таковы, в частности, отдельные водные обитатели с наружным способом оплодотворения, при котором нет необходимости в непосредственной встрече партнеров, например одиночные актинии. У видов с внутренним оплодотворением встречи самцов и самок также могут быть очень кратковременными, лишь для осуществления копуляции, в остальное время животные живут независимо друг от друга. Таков образ жизни многих насекомых, например божьей коровки, хищных жуков-жужелиц и др.
У видов с одиночным образом жизни часто образуются временные скопления особей – в местах зимовок, в период, предшествующий размножению, и т. п. Так, бабочки– крапивницы поздней осенью целыми гроздьями собираются в чердачных помещениях или других укрытиях, жужелицы и божьи коровки – в сухой подстилке возле пней и комлей деревьев, сомы и щуки – в зимовальных ямах на дне водоема. У бентосных животных такие скопления могут быть постоянными, например устричные или мидиевые банки, агрегации полихет или поселения морских желудей. В этих поселениях увеличиваются шансы на возможность оплодотворения половыми продуктами, выбрасываемыми в воду. Однако подобные агрегации не сопровождаются установлением более или менее закономерного поведения животных по отношению друг к другу, и каждое из них относительно независимо от остальных.
Дальнейшее усложнение отношений внутри популяции осуществляется по двум направлениям: усилению связи между половыми партнерами и возникновению контактов между родительским и дочерним поколениями. На этой основе в популяциях формируются семьи, которые очень разнообразны по составу и длительности существования.
Родительские пары могут создаваться как на короткий, так и на длительный срок, а у некоторых видов – на всю жизнь взрослых особей. Например, среди птиц полигамные тетерева, глухари спариваются со многими самками, не образуя устойчивых семейных пар. У ряда уток – кряквы, шилохвости и др. – пары подбираются еще на местах зимовок или во время пролета, но, после того как самка садится на гнездо, самец ее покидает. У большинства воробьиных самец и самка держатся вместе в течение всего периода гнездования. Семейные пары лебедей, журавлей, голубей сохраняются на долгие годы.
Выбор партнеров сопровождается у животных особым брачным поведением, часто достигающим большой сложности. У пауков–скакунчиков самцы добиваются спаривания с самкой после своеобразного танца, в продолжение которого они описывают возле нее суживающиеся полукруги, принимая особые позы, а приблизившись, начинают быстро вертеться, увлекая самку. Считают, что предкопуляционное поведение самцов пауков тормозит инстинкт агрессии у самок, которые иначе реагируют на них как на добычу. Дальнейшее поведение пауков различно: у многих всеядных видов самец после введения им спермы в половые пути самки становится ее жертвой, у бегающих Lycosidae – поспешно спасается бегством, у большинства скакунчиков Salticidae самец и самка расходятся спокойно, а у ряда видов семейства Dictinidae некоторое время мирно уживаются в одном гнезде.
Широко известное «ухаживание» у птиц и млекопитающих также предотвращает агрессивные и оборонительные реакции особей противоположного пола, приводит к синхронизации процессов полового созревания и стимулирует у партнеров готовность к спариванию (рис. 87). Это имеет большое значение для осуществления размножения, к которому самец и самка должны быть готовы в одно и то же время.

Рисунок 87 – Элементы брачного поведения обыкновенной чайки (по Н. Тинбергену, 1970)
Выбор половых партнеров часто сопровождается усилением конкурентных отношений в популяциях животных. В период, предшествующий размножению, у многих видов возникают драки самцов, ритуальные демонстрации и другие типы специализированного поведения, направленного на устранение конкурентов. Эти столкновения, несмотря на частую ожесточенность, редко приводят к серьезным ранениям соперников, обычно ограничиваясь изгнанием одного из них с территории, где находятся самки. У рыб, например, в таких сражениях преобладает ритуал угрозы или противники кусают друг друга в пасть – наименее уязвимую часть тела, но никогда не наносят более опасных укусов в бок. Токующие турухтаны стремительно бросаются друг на друга, но затем внезапно замирают с взъерошенным «воротником», после чего вновь повторяют броски.
Таким образом, период, предшествующий размножению в популяциях животных, характеризуется активным поиском и резким усилением контактов между особями.
Семейный образ жизни. При семейном образе жизни усиливаются также связи между родителями и их потомством. Простейший вид такой связи – забота одного из родителей об отложенных яйцах: охрана кладки, инкубация, дополнительное аэрирование и т. п. Так, среди земноводных мраморная амбистома откладывает яйца в ямки во влажной лесной подстилке и остается их сторожить. Самка темной саламандры обвивает кладку своим телом и не покидает ее до выклева личинок. Самец трехиглой колюшки после охраны кладки и заботы о ней некоторое время оберегает и мальков. У птиц сложные заботы о птенцах продолжаются до поднятия их на крыло, а у ряда крупных млекопитающих, например у медведей, тигров, детеныши воспитываются в семейных группах в течение нескольких лет, до наступления их половой зрелости.
Различают семьи отцовского, материнского и смешанного типа, в зависимости от того, кто из родителей берет на себя уход за потомством. В семьях с устойчивым образованием пар в охране и выкармливании молодняка принимают обычно участие и самец и самка.
При семейном образе жизни территориальное поведение животных выражено наиболее ярко: различные сигналы, маркировка, ритуальные формы угрозы и прямая агрессия обеспечивают владение участком, достаточным для выкармливания потомства.
Более крупные объединения животных – стаи, стада и колонии. В основе их формирования лежит дальнейшее усложнение поведенческих связей в популяциях.
Колонии. Это групповые поселения оседлых животных. Они могут существовать длительно или возникать лишь на период размножения, как, например, у многих птиц – грачей, чаек, гагар, тупиков и т. п. По сложности взаимосвязей между особями колонии животных чрезвычайно разнообразны – от простых территориальных скоплений одиночных форм до объединений, где отдельные члены выполняют, как органы в целостном организме, разные функции видовой жизни.
Поселения морских желудей, например, можно рассматривать и как агрегации одиночных особей, и как элементарную форму колонии, так как одновидовые скопления возникают у них не случайно, а на основе хемотаксиса, свойственного личинкам. Взаимоотношения между особями в таких примитивных объединениях сводятся к стимуляции синхронного полового созревания химическими выделениями членов колонии, а конкуренция за облавливаемое пространство в известной мере предотвращается ориентировочной реакцией личинок перед прикреплением.
Более сложная форма колонии – такие поселения животных, в которых некоторые функции их жизни выполняются сообща, что увеличивает вероятность выживания отдельных особей. Чаще всего такими общими функциями становятся защита от врагов и предупредительная сигнализация. Чайки, кайры, некоторые гуси, ласточки и другие птицы обычно с шумом набрасываются на хищника, угрожающего птенцам или кладкам. Тревога, поднятая любой заметившей опасность птицей, мобилизует остальных. Сообща птицам удается изгонять даже крупных хищников, с которыми они не справились бы поодиночке, – песцов, ястребов, сов и др.
В колониальных поселениях птиц часто сохраняются индивидуальные гнездовые участки. У серебристых чаек, например, колонии рыхлые – между гнездами сохраняется расстояние в 3–5 м. При выборе участков затеваются драки. В поселениях некоторых других птиц подобные территориальные инстинкты не проявляются совсем. Например, городские ласточки лепят свои гнезда часто вплотную одно к другому.
Общественные ткачики (Philacterus socius) строят на деревьях большое общее гнездо из травы с многочисленными отверстиями, ведущими в индивидуальные гнездовые полости. Южноамериканские кукушки–личинкоеды ани строят совместные гнезда, в которые откладывают яйца около десятка самок. Эту общую кладку насиживают одновременно несколько птиц, периодически сменяемые другими.
Среди млекопитающих колониальны сурки, вискачи, пищухи, пеструшки. Колонии млекопитающих чаще возникают не как территориальные объединения разных семей, а на основе разрастания семейных групп, с сохранением связей между отпочковывающимися семьями. Зверьков из других колоний при полной занятости территории изгоняют, но в период низкой численности популяции они могут вливаться в состав поселения, которое, таким образом, приобретает смешанный характер.
Наиболее сложные колонии у общественных насекомых – термитов, муравьев, пчел. Они возникают на основе сильно разрастающейся семьи. В таких колониях–семьях насекомые выполняют сообща большинство основных функций: размножения, защиты, обеспечения кормом себя и потомства, строительства и т. п. При этом существует обязательное разделение труда и специализация отдельных особей или возрастных групп на выполнении определенных операций. Члены колонии действуют на основе постоянного обмена информацией друг с другом.
У рыжих лесных муравьев Formica rufa колонии могут занимать одно или несколько гнезд, связанных между собой системой дорог. Самцы выполняют только функцию оплодотворения самок, после чего погибают. Самки сначала осуществляют расселение, затем основывают новую семью, выполняя все операции, связанные с воспитанием первого потомства. После этого они специализируются только на производстве яиц, а остальную работу в муравейнике выполняют рабочие – особая каста, представленная бесплодными измененными самками. Среди них также существует разделение труда. Определенные группы обеспечивают сбор строительного материала, уход за тлями и сбор пади, охоту и разведку, транспортировку добычи, обмен молодью между гнездами и т. п. Кроме того, у рабочих муравьев существует и возрастная смена функций. В течение 1–1,5 месяца жизни они работают внутри гнезда, ухаживают за молодью, а позднее становятся строителями, разведчиками, фуражирами.
Среди муравьев одной колонии постоянно осуществляется обмен пищей. Муравьи кормят друг друга содержимым зобика, желудка, слизывают выделения различных желез. Это явление получило название трофаллаксиса. На основе трофаллаксиса объединение семьи поддерживается не только общим пищевым потоком, но и действием различных биологически активных веществ, содержащихся в выделениях.
В организации совместной деятельности у муравьев большую роль играет достаточно разнообразная система сигнализации: позами, движениями усиков, прикосновениями разного характера, пахучими выделениями и т. п.
Таким образом, по мере усложнения колониального объединения поведение, а иногда также физиология и строение отдельной особи все больше подчиняются интересам всей колонии.
Особые формы колониальности развиваются у животных с вегетативным размножением, в частности у кишечнополостных и некоторых других водных обитателей. Разделение функций у полипов, связанных между собой системой распределительных каналов, заходит у некоторых видов настолько далеко, что вся колония выглядит и ведет себя как единая особь. Наиболее наглядный пример – сифонофоры, в строении которых практически стираются границы между колонией и цельным организмом, имеющим систему дифференцированных органов. Образование таких колоний-организмов происходит на основе единого метаболизма.
Стаи. Это временные объединения животных, которые проявляют биологически полезную организованность действий. Стаи облегчают выполнение каких-либо функций в жизни вида: защиты от врагов, добычи пищи, миграции. Наиболее широко стайность распространена среди птиц и рыб, у млекопитающих характерна для многих собачьих. В стаях сильно развиты подражательные реакции и ориентация на соседей.
По способам координации действий стаи делятся на две категории: - эквипотенциальные, без выраженного доминирования отдельных членов и
- стаи с лидерами, в которых животные ориентируются на поведение одной или нескольких, обычно наиболее опытных, особей.
Объединения первого типа характерны в основном для рыб, но известны также у мелких птиц, перелетной саранчи и некоторых других форм. Второй тип стай встречается обычно у крупных птиц и млекопитающих.
Стаи рыб очень изменчивы по величине, форме, плотности. Они часто переформировываются, иногда по нескольку раз в сутки. Обычно рыбы группируются в стаи лишь в светлое время суток, при зрительном контакте с другими особями, и рассредоточиваются на ночь. Защитная роль стайных объединений рыб очень велика. В опытах одиночные рыбки вылавливаются хищником в несколько раз быстрее, чем члены стаи. В группе осуществляется «круговой обзор», благодаря которому хищнику труднее приблизиться незамеченным. Кроме того, многочисленность двигающихся особей дезориентирует врага. Тела рыб, непрерывно меняющих направление движения, создают мерцание, затрудняющее фиксацию взгляда на отдельных особях, и делают невозможными целенаправленные броски. Стая быстро маневрирует при опасности, обтекая хищника, который, бросившись в ее середину, оказывается в пустоте (рис. 88). Для поведения рыб в стае характерен имитационный рефлекс – подражание действиям соседей.
У птиц стаи формируются при сезонных перелетах или, у оседлых и кочующих форм, при зимних кормежках. Стаи при перелетах образуют те виды, которым свойственно колониальное гнездование или коллективное кормление. Одиночно гнездящиеся и кормящиеся виды стай в полете не образуют.

Рисунок 88 – Маневр стаи верховки при броске щуки (по Д. В. Радакову, 1972)
В стаях оседлых птиц существует постоянная сигнализация, звуковая и зрительная связь между особями. Благодаря внутристайной, а часто и межстайной сигнализации птицы используют и опыт, и случайное обнаружение отдельными особями благоприятных мест ночевок и отдыха, источников корма, восходящих потоков воздуха и т. п.
Волчьи стаи возникают для групповой охоты зимой. В группе зверям удается справиться с крупными копытными, охота на которых в одиночку безрезультатна. При групповой охоте волков часто практикуется преследование с выходом на перехват жертвы, нагон жертвы на засаду или захват ее в кольцо, что требует согласованности и координации действий всех особей. Стайность известна для гиен, гиеновых собак, койотов и др. В стаях млекопитающих велика роль вожаков и специфичны отношения между отдельными особями, что сближает эти групповые образования со стадами.
Стада. Это более длительные и постоянные объединения животных по сравнению со стаями. В стадных группах, как правило, осуществляются все основные функции жизни вида: добывание корма, защита от хищников, миграции, размножение, воспитание молодняка и т. п. Основу группового поведения животных в стадах составляют взаимоотношения доминирования–подчинения, основанные на индивидуальных различиях между особями.
Один из вариантов организации стад – это группы с временными либо относительно постоянными лидерами – особями, на которых концентрируется внимание других и которые своим поведением определяют направление перемещения, места кормежки, реакцию на хищников и другие свойства стада. Стадо действует как единое целое, подражая лидеру. Деятельность лидера не направлена непосредственно на подчинение других особей. Лидером становится более опытный член стада. Например, стада северных оленей обычно ведут старые важенки. Они лучше других ориентируются при миграциях и нападении хищников, так как периодически им приходится делать это в одиночку. Перед рождением олененка самка обычно удаляется и несколько дней, пока не окрепнет малыш, вынуждена одна оберегать и защищать его, а затем вместе с ним догоняет стадо. При переменном лидерстве одни животные становятся во главе стада чаще других. Большое значение имеют при этом также особенности высшей нервной деятельности. В стадах северных оленей около 18–20 % животных являются потенциальными лидерами. У других животных лидеры могут быть более постоянны.
Биологическое значение лидерства в том, что индивидуальный опыт отдельных особей может быть использован всей группой.
В крупных стадах выделяются также объединения, представляющие семейные или возрастные группы, внутри которых контакты более дружелюбны, чем с членами других аналогичных групп. Во внутристадных группировках, независимо от общего лидерства, могут складываться отношения доминирования-подчинения.
Наиболее сложна поведенческая организация стад с вожаками и иерархическим соподчинением особей. Вожаки, в отличие от лидеров, характеризуются поведением, непосредственно направленным на активное руководство стадом: специальными сигналами, угрозами и прямым нападением. В подобных стадах часто возникают разделение «прав» и «обязанностей» и более сложные формы общественного поведения, выгодные для групп в целом.
Ранг каждой особи в стаде определяется многими причинами. Имеют значение возраст, физическая сила, опыт и наследственные качества животного. Более сильные и опытные, с устойчивым типом нервной системы, как правило, доминируют над более слабыми. Доминирование проявляется в преимуществе при поедании пищи, праве на самку, передвижении в группе и т. п.
Системы доминирования–подчинения очень различны у разных видов. При линейной иерархии в ряду рангов А–В–С и т. д. особи, принадлежащие к каждому рангу, подчинены предыдущим, но главенствуют над последующими. Последние в таком ряду животные – самые бесправные в группе. Подобные отношения возникают, например, в стаях ездовых собак, где вожаки активно подчиняют себе стаю, угрожая и задавая трепку непослушным. Животные низшего ранга проявляют признаки покорности перед всеми остальными, подходят к пище в последнюю очередь, их не подпускают к самкам, изгоняют с лучших мест отдыха и т. д. Иерархия в группах часто бывает более сложной. Например, могут возникать параллельные ряды подчинения: один среди самцов, другой – среди самок, как, например, у некоторых игрунковых обезьян. У павианов, помимо того, самцы доминируют над самками, а те в свою очередь над детенышами. У некоторых видов обнаружено иерархическое соподчинение по типу «треугольника»: А нападает на В, В – на С, а С подчиняет себе А. Такое соотношение может сохраняться в группе довольно долго. Одним из вариантов иерархии может быть деспотия – доминирование одного животного над всеми остальными членами группы.
Ранг животного в группе определяется столкновениями между особями, которые могут иметь характер прямой борьбы или ритуальных угроз. После установления ранга всех членов группы прямые столкновения между ними прекращаются и порядок поддерживается сигнальным или ритуальным поведением.
Иерархически организованному стаду свойствен закономерный порядок перемещения, определенная организация при защите, расположении на местах отдыха и т. п. Так, при передвижении семейного стада зебр впереди всегда идет старая кобыла, за ней следуют сначала самые молодые, а затем старшие жеребята, за которыми двигаются взрослые зебры. Шествие завершает жеребец-вожак. В стаде павианов в центре, в наибольшей безопасности, находятся самки с детенышами или готовящиеся к размножению, по краям – вожаки, молодые самцы и неразмножающиеся самки (рис. 89). Впереди и позади стада шествуют крупные самцы, готовые отразить нападение. Описаны случаи, когда при преследовании стада хищниками самец–доминант возвращался за отставшим детенышем, несмотря на грозившую ему большую опасность.
Вожаки-доминанты выполняют разнообразные общественные функции. В табуне лошадей, например, вожак управляет движением, уводит косяк от опасности, защищает от хищников, прекращает драки, проявляет заботу о жеребятах и больных животных и т. п. В сложной обстановке вожаки ряда видов (лошади Пржевальского, павианы) могут проводить разведку, отделяясь от стада, а затем возвращаясь к нему и определяя безопасное направление движения.
Конфликты в группах, имеющих вожака, обычно решаются с его участием или в его присутствии, у разных видов с разной степенью агрессивности. В стадах павианов доминанты могут прибегать к преследованию и укусам, тогда как у горилл вожак нередко восстанавливает порядок лишь взглядом или движением головы.
Биологический смысл иерархической системы доминирования-подчинения заключается в создании согласованного поведения группы, выгодного для всех ее членов. После «расстановки сил» животные не тратят лишней энергии на индивидуальные конфликты, а группа в целом получает преимущества, подчиняясь наиболее сильным и опытным индивидуума. Согласованность поведения имеет большое значение в обеспечении защиты от хищников, предупреждении опасности, выращивании молодняка, миграциях и т. п. В сложных условиях, например при голодовках, гибнут в первую очередь более слабые, подчиненные особи, но под защитой группы они все же имеют больше шансов выжить, чем в одиночку.
Иерархия в групповых отношениях адаптивна прежде всего потому, что она динамична. В жизни стада идет непрерывная проверка соответствия его структуры меняющимся условиям. Состав группы также не остается постоянным: взрослеет молодняк, появляются новые пришельцы, старые животные теряют силы, часть особей погибает. Эти изменения сопровождаются перестройкой рангов. Переходу особи в новый ранг обычно предшествуют прямые или ритуальные схватки и демонстрации, «пересмотр сил», после чего закрепляется новое положение. На роль вожака–доминанта всегда претендуют более молодые, набирающие силу и опыт животные. Они занимают его место после первого же проявления вожаком неспособности отстоять свои права.
Иерархия наиболее ярко выражена в стадах, но она характерна и для многих групп оседлых животных – семей, колоний. Иерархия может проявляться также в распределении территориальных участков у одиночных форм. Среди сусликов, охраняющих свои индивидуальные территории, доминирующие особи занимают наиболее благоприятные места в поселениях. Среди колониальных птиц (чаек, гусей, цапель и др.) наиболее крупные и опытные гнездятся в центре колонии, тогда как периферию занимают физиологически менее полноценные особи, проявляющие реакцию подчинения по отношению к доминантам.
Система доминирования–подчинения выявлена и у ряда беспозвоночных: насекомых (сверчков, жуков–чернотелок и др.), некоторых ракообразных и пр.
Установление иерархии сопровождается изменениями в гормональной системе организмов. У млекопитающих, например, возникает гипертрофия коры надпочечников. Эти изменения оказываются наибольшими у животных, находящихся в самом низу иерархической лестницы, и не проявляются у доминантов. Стероидные гормоны, выделяемые корой надпочечников, оказывают большое влияние на процессы обмена веществ, которые, в свою очередь, лежат в основе изменения поведения.
Эффект группы. Жизнь в группе через нервную и гормональную системы отражается на протекании многих физиологических процессов в организме животного. У изолированных особей заметно меняется уровень метаболизма, быстрее тратятся резервные вещества, не проявляется целый ряд инстинктов и ухудшается общая жизнеспособность.
Оптимизация физиологических процессов, ведущая к повышению жизнеспособности при совместном существовании, получила название эффект группы. Он проявляется как психофизиологическая реакция отдельной особи на присутствие других особей своего вида. У овец вне стада учащаются пульс и дыхание, а при виде приближающегося стада эти процессы нормализуются. Одиночно зимующие летучие мыши отличаются более высоким уровнем обмена веществ, чем в колонии. Это ведет к повышенной трате энергии, истощению и часто заканчивается гибелью животных.
Затраты организмом энергии на основную жизнедеятельность учитывают, измеряя скорость потребления кислорода животными в покое. Эти показатели возрастают у всех животных, приспособленных к групповой жизни и оказавшихся вне стада, семьи или колонии. Напротив, даже простое скучивание сокращает газообмен порой до 50 %. Возможно, что это явление лежит в основе агрегации на зимовку рыб, лягушек, моллюсков, насекомых, у которых также обнаружена экономия энергетических затрат в скоплениях.
Эффект группы проявляется в ускорении темпов роста животных, повышении плодовитости, более быстром образовании условных рефлексов, повышении средней продолжительности жизни индивидуума и т. д. В группе животные часто способны поддерживать оптимальную температуру (при скучивании, в гнездах, в ульях). У многих животных вне группы не реализуется плодовитость. Голуби некоторых пород не откладывают яйца, если не видят других птиц. Достаточно поставить перед самкой зеркало, чтобы она приступила к яйцекладке. Кайры при небольшой численности пар в колонии начинают размножение только тогда, когда колония увеличится по периферии за счет других видов: чистиков, тупиков, моевок.
Эффект группы не проявляется у видов, ведущих одиночный образ жизни. Если таких животных искусственно заставить жить вместе, у них повышается раздражительность, учащаются столкновения и многие физиологические показатели сильно уклоняются от оптимума, о чем можно судить, например, по увеличению энергетических затрат на основные процессы жизнедеятельности. Так, ушастые ежи в группе повышают потребление кислорода до 134 % по сравнению с содержащимися поодиночке.
Положительный эффект группы проявляется лишь до некоторого оптимального уровня плотности популяции. Если животных становится слишком много, это грозит для всех недостатком ресурсов среды. Тогда вступают в действие другие механизмы, приводящие к снижению численности особей в группе путем ее деления, рассредоточения или падения рождаемости.
Группа закономерностей, характерных для пространственных изменений особей в популяциях, сменяющих друг друга при движении с севера на юг (или наоборот) и живущих в различных условиях островной изоляции, наиболее известна. Кажется, еще Фридрих I Барбаросса (1125–1190) знал правило измельчания размеров тела теплокровных животных при движении с севера на юг, названное затем правилом К. Бергмана (1847): у теплокровных животных, подверженных географической изменчивости, размеры тела особей статистически (в среднем) больше у популяций, живущих в более холодных частях ареала вида. Правилу Бергмана подчиняются около 50 % видов теплокровных животных, подавляющая часть которых (до 90 %) птицы. Механизм действия правила Бергмана очевиден – укрупнение размеров особей термодинамически позитивно меняет отношение объема и площади тела, снижает уровень основного обмена. В то же самое время правило не может действовать в тех случаях, когда наблюдается недостаток кормов или колеблется их доступность: при увеличении размеров тела общая масса потребляемого корма возрастает.
Как бы дополнением к правилу Бергмана служит правило Р. Гессе: особи популяций в северных районах обладают относительно большей массой сердца (при ее сравнении с массой тела), чем особи популяций, живущих в южных районах или в более теплых местообитаниях. Большая интенсивность обмена и необходимость поддерживать относительно постоянную температуру тела при более низких температурах требует лучшего кровоснабжения. А для него нужна более мощная сердечная мышца.
Столь же очевидны причины действия закона покровов (покрытия) тела: плотность (густота и терморегулирующая способность) покровов тела млекопитающих и птиц достигает максимума в холодных и засушливых областях. Это помогает животным противостоять экстремальным температуре и влажности. Уточнение к закону покровов (покрытия) тела служит правило густоты мехового покрова: у млекопитающих холодных местообитаний мех как правило гуще, чем у живущих в теплых географических зонах и поясах. Часто это правило справедливо и для млекопитающих сухого климата, где густой мех предохраняет от излишнего испарения воды с поверхности тела, а также от перегревания.
Как бы частным случаем правила Бергмана, а скорее, более общего правила поверхностей служит правило Д. Аллена (1877): увеличение выступающих частей тела одного вида теплокровных животных (конечностей, хвоста, ушей и т. п.) при продвижении с севера на юг. Достаточно ясно, что удлинение выступающих частей тела увеличивает поверхность тела и его теплоотдачу.
Для рыб существует особое правило числа позвонков, или правило А. Жордана, согласно которому (у сельдей, трески и др.) в водоемах с повышенной соленостью и более низкими температурами в хвостовой части тела возрастает число позвонков. Правило Жордана не имеет отношения к правилу Аллена как таковому, а связано с приспособлением к движению в более плотной среде. Подобного рода закономерностей много у почвенных и глубоководных организмов – их удлиненность, часто червеобразность и т. п.
Климатические особенности воздействуют и на цвет особей популяций, живущих на севере и на юге, в сухих и влажных местообитаниях. Эту специфику отражает правило, или закон К. Глогера (1833): виды животных северных и влажных географических зон имеют более интенсивную окраску, чем обитатели южных сухих регионов. Темная окраска, отражая меньше тепла, улучшает его аккумуляцию телом животного. В то же самое время многие виды млекопитающих и птиц Севера имеют светлую окраску, что объясняется не столько самим цветом мехового покрова, сколько его структурой, строением волос, одновременно отражающих свет и работающих как кумулирующие линзы. Кроме того, на льду белый цвет служит и защитной окраской (например, у бельков тюленей).
Многие признаки островных популяций видов отличаются от имеющихся у материковых. Объясняет это явление принцип основателя: единичные особи-основатели новой (изолированной) колонии или популяции несут в себе лишь часть общей генетической информации, присущей виду. Нехватка потенциала наследственной изменчивости может лежать в основе правила островного измельчания: популяции видов животных, обитающие на островах, как правило, образуются более мелкими особями, чем материковые, живущие в аналогичных условиях. Правило островного измельчания имеет много исключений, особенно на крупных островах (например, кабарга Сахалина крупнее чем материковая), но, видимо, статистически верно.
Половая структура. В зависимости от способа размножения различают популяции:
- панмиктические;
- клональные;
- клонально-панмиктические.
Панмиктическими называют популяции, состоящие из особей, размножающихся половым путем, для которых характерно перекрестное оплодотворение.
Клональными называют популяции, состоящие из особей, для которых характерно только бесполое размножение.
В том случае, когда половое размножение сочетается с бесполым, говорят о клонально–панмиктических популяциях.
Половая структура популяции по генетическим законам должна быть представлена равным соотношением мужских и женских особей, то есть 1:1. Но в силу специфики физиологии и экологии, свойственной разным полам, в силу их разной жизнеспособности, влияния факторов внешней среды, социальных, антропогенных могут быть значительные различия в этом соотношении. И эти различия неодинаковы как в разных популяциях, так и в разных возрастных группах одной и той же популяции. Это наглядно показано на рисунке 8, представляющим срезы возрастной и половой структуры для населения бывшего СССР и африканской республики Кения. На срезе СССР, на фоне естественного распределения возрастных групп в жизненном цикле, очевидно снижение рождаемости в годы войны и увеличение ее в послевоенные годы. Диспропорция между женским и мужским полом тоже, несомненно, связано с войной. В Кении же прослеживается закономерная связь распределения полов и явной убыли населения в предрепродуктивном возрасте с низким уровнем жизни, зависимостью от природных условий.
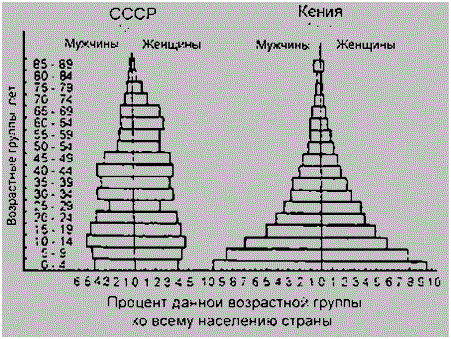
Рисунок 89 – Половозрастные пирамиды населения бывшего СССР и Кении (по Н.Ф. Реймерсу, 1990)
Различают первичное, вторичное и третичное соотношения полов. Первичное соотношение – соотношение, наблюдаемое при формировании половых клеток (гамет). Обычно оно равно 1:1. Такое соотношение обусловлено генетическим механизмом определения пола. Вторичное соотношение – соотношение, наблюдаемое при рождении. Третичное соотношение – соотношение, наблюдаемое у взрослых половозрелых особей. Например, у человека во вторичном соотношении несколько преобладают мальчики, в третичном – женщины: на 100 мальчиков рождается 106 девочек, к 16–18 годам из–за повышенной мужской смертности это соотношение выравнивается и к 50 годам составляет 85 мужчин на 100 женщин, а к 80 годам – 50 мужчин на 100 женщин.
У некоторых рыб (р. Пецилия) различают три типа половых хромосом: Y, Х и W, из них Y–хромосома несет гены мужского пола, а Х и W–хромосомы – гены женского пола, но разной степени «мощности». Если генотип особи имеет вид YY, то развиваются самцы, если XY – самки, если же WY, то в зависимости от условий среды развиваются половые признаки самца или самки.
Изучение половой структуры популяций очень важно, поскольку между особями разных полов сильно выражены как экологические, так и поведенческие различия. Например, сильно различаются между собой самцы и самки комаров (сем. Culicidae): по темпам роста, срокам полового созревания, устойчивости к изменению температуры. Самцы в стадии имаго не питаются совсем или питаются нектаром, а самкам необходимо напиться крови для полноценного оплодотворения яиц. У некоторых видов мух популяции состоят только из самок.
Даже если образ жизни самцов и самок сходен, они различаются по многим физиологическим признакам: темпам роста, срокам полового созревания, устойчивостью к изменениям температуры, голоданию и т. п.
Различия в смертности проявляются еще в эмбриональный период. Например, у ондатр во многих районах среди новорожденных в полтора раза больше самок, чем самцов. В популяциях пингвинов Megadyptes antipodes при выходе птенцов из яиц подобной разницы не отмечается, но к десятилетнему возрасту на каждых двух самцов остается лишь одна самка. У некоторых летучих мышей доля самок в популяции после зимней спячки снижается порой до 20 %. Многие другие виды отличаются, наоборот, более высокой смертностью самцов (фазаны, утки-кряквы, большие синицы, многие грызуны).
У рыжих лесных муравьев (Formica rufa) из яиц, отложенных при температуре ниже +20 0C, развиваются самцы, при более высокой – почти исключительно самки. Механизм этого явления заключается в том, что мускулатура семяприемника, где хранится после копуляции сперма, активизируется лишь при высоких температурах, обеспечивая оплодотворение откладываемых яиц. Из неоплодотворенных же яиц у перепончатокрылых развиваются лишь самцы.
Хорошо прослеживается роль экологических факторов в формировании половой структуры у видов с чередованием половых и партеногенетических поколений. При оптимальной температуре у дафнии (Daphnia magna) популяцию образуют партеногенетические самки, а при отклонениях от нее - появляются и самцы. В популяциях меченосцев соотношение полов зависит от значения рН среды. При рН–6,2 количество самцов в потомстве составляет 87–100 %, а при рН–7,8 – от 0 до 15%.
Есть виды, у которых пол изначально определяется не генетическими, а экологическими факторами, как, например, у Ариземы японской при образовании массы клубней женские соцветия формируются на растениях с крупными мясистыми клубнями, а мужские – на растениях с мелкими.
Среди цветковых растений встречается немало двудомных видов, у которых существуют мужские и женские особи: виды ив, тополей, дрема белая, щавель малый, пролесник многолетний, бодяк полевой и др. Есть и виды с женской двудомностью, когда одни особи имеют обоеполые цветки, а другие – женские, т. е. с неразвитым андроцеем. Обычно андростерильные цветки мельче, чем обоеполые. Такое явление встречается в семействах губоцветных, гвоздичных, ворсянковых, колокольчиковых и др. Примерами видов с женской двудомностью являются чабрец Маршалла, душица обыкновенная, мята полевая, будра плющевидная, смолевка поникшая, герань лесная и др. Популяции таких видов генетически неоднородны. В них облегчено перекрестное опыление, чаще наблюдается протероандрия – более раннее созревание пыльников по сравнению с пестиками. В пределах ареала видов половая структура популяций растений более или менее постоянная, однако изменение внешних условий меняет соотношение полов. Так, в засушливый 1975 г. в Зауралье резко уменьшилось число женских форм, например у шалфея степного в 10 раз, у спаржи лекарственной в 3 раза.
Таким образом, соотношение полов в популяции устанавливается не только по генетическим законам, но и в известной мере под влиянием среды.
Возрастная структура – характеризуется соотношением особей по возрастам, фазам развития. Возрастная структура влияет на рождаемость, смертность, определяет способность к размножению в данный момент и дает возможность оценить перспективы популяции.
Возрастная структура также отражает скорость обновления популяции и взаимодействие возрастных групп с внешней средой. Она зависит от особенностей жизненного цикла, существенно различающегося у разных видов (например, птиц и у млекопитающих хищников), и внешних условий.
На соотношения возрастных групп влияют:
- общая продолжительность жизни;
- время достижения половой зрелости;
- интенсивность размножения;
- степень приспособления к определенным условиям.
В жизненном цикле особей обычно выделяют три возрастных периода: предрепродуктивный (ювенильный), репродуктивный и пострепродуктивный (синильный) . Для растений характерен еще период первичного покоя, который они проходят в стадии поящихся семян. Каждый из периодов может быть представлен одной (простая структура) или несколькими (сложная структура) возрастными стадиями. Простой возрастной структурой обладают однолетние растения, многие насекомые. Сложная структура характерна для разновозрастных популяций деревьев, для высокоорганизованных животных. Чем сложнее структура, тем выше приспособительные возможности популяции.
В геоботанике получила признание классификация растений по возрасту Н.М. Черновой, А.М. Быловой:
- покоящиеся семена;
- проростки (всходы) – растения первого года жизни, многие из них живут за счет питательных веществ в семядолях;
- ювенильные – переходят к самостоятельному питанию, но размерами и морфологически еще отличаются от взрослых растений;
- имматурные – обладают переходными признаками от ювенильных к взрослым растениям, еще очень малы, у них идет смена типа нарастания, начинается ветвление побегов;
- виргинильные – «взрослые подростки», могут достигать размеров взрослых особей, но регенеративные органы отсутствуют;
- молодые генеративные – характерно наличие генеративных органов, завершается формирование облика, типичного для взрослого растения;
- средневозрастные генеративные – отличаются максимальным годичным приростом и максимальной репродуктивностью;
- старые генеративные – растения продолжают плодоносить, но у них полностью прекращаются рост побегов и образование корней;
- субсенильные – плодоносят очень слабо, идет отмирание вегетативных органов, новообразование побегов идет за счет спящих почек;
- сенильные – очень старые, дряхлые особи, появляются черты ювенильных растений: крупные одиночные листья, порослевые побеги.
Ценопопуляция, в которой представлены все перечисленные стадии, называется нормальной полночленной.
В лесоведении и таксации принята классификация древостоев и насаждений по классам возраста. Для хвойных пород:
- проростки и самосев – 1–10 лет, высота до 25 см;
- стадия молодняка – 10–40 лет, высота от 25 до 5 м; под пологом леса соответствует мелкому (до 0,7 м), среднему (0,7–1,5 м) и крупномерному (>1,5 м) подросту;
- стадия жердняка – средневозрастные насаждения 50–60 лет; диаметры стволов от 5 до 10 см, высота – до 6–8 м; под пологом леса молодое поколение древостоя, или тонкомер с аналогичными размерами;
- приспевающие насаждения – 80–100 лет; по размерам могут незначительно уступать материнском деревьям, на открытом месте и в редколесьях обильно плодоносят; в лесу могут еще находиться во втором ярусе, не плодоносят; ни в коем случае не назначаются в рубку;
- спелые древостои – 120 лет и старше, деревья первого яруса и отставшие в росте деревья второго яруса; обильно плодоносят, в начале этой стадии достигают технической спелости, в конце – биологической;
- перестойные – старше 180 лет, продолжают обильно плодоносить, но постепенно дряхлеют и усыхают или вываливаются еще будучи живыми.
Для лиственных пород градации по размерам аналогичные, но в связи с их более быстрым ростом и старением класс возраст у них составляет не 20, а 10 лет.
Соотношение возрастных групп в структуре популяции характеризуют ее способность к размножению и выживанию, и согласуется с показателями рождаемости и смертности. В растущих популяциях с высокой рождаемостью преобладают молодые (рис. 90), еще не репродуктивные особи, в стабильных – обычно это разновозрастные, полночленные популяции, у которых регулярно определенное число особей переходит из младших возрастных групп в старшие, рождаемость равна убыванию населения. В сокращающихся популяциях основу составляют старые особи, возобновление в них отсутствует или совсем незначительно.
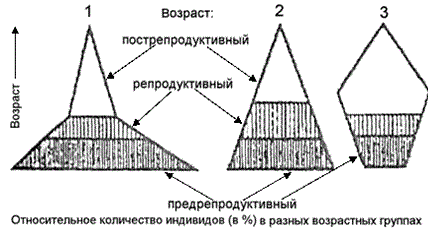
Рисунок 90 – Типы популяций: 1 – растущая (поползень); 2 – стабильная (барсук); 3 – сокращающаяся (тигр амурский)
Популяция приспосабливается к изменению условий среды путем обновления и замещения особей, то есть процессами рождения (возобновления) и убывания (отмирания), дополняемыми процессами миграции. В стабильной популяции темпы рождаемости и смертности близки, сбалансированы. Они могут быть непостоянны, но плотность популяции незначительно отличается от какой–то средней величины. Ареал вида при этом ни увеличивается, ни уменьшается.
В растущей популяции рождаемость превышает смертность. Для растущих популяций характерны вспышки массового размножения, особенно у мелких животных (саранча, 28–точечная картофельная коровка, колорадский жук, грызуны, вороны, воробьи); из растений (амброзия, борщевик Сосновского в северной республике Коми, одуванчик, прилипало гималайское, отчасти – дуб монгольский). Нередко растущими становятся популяции крупных животных в условиях заповедного режима (лоси в Магаданском заповеднике, на Аляске, олень пятнистый в Уссурийском заповеднике, слоны в национальном парке Кении) или интродукции (лось в Ленинградской области, ондатра в Восточной Европе). При переуплотнении у растений (обычно совпадает с началом сомкнутости покрова, кронового полога) начинается дифференциация особей по размерам и жизненному состоянию, самоизреживание популяций, а у животных (обычно совпадает с достижением половой зрелости молодняка) начинается миграция на сопредельные свободные участки.
Если смертность превышает рождаемость, то такая популяция считается сокращающейся. В естественной среде она сокращается до определенного предела, а затем рождаемость (плодовитость) вновь повышается и популяция из сокращающейся становится растущей. Чаще всего неумеренно растущими бывают популяции нежелательных видов, сокращающимися – редких, реликтовых, ценных, как в экономическом, так и в эстетическом отношении.
Наличие сложной половозрастной структуры обусловливает внутрипопуляционную дифференциацию экологических ниш, особенно, у животных с полным метаморфозом. Экологическая ниша всей популяции распадается на субниши, которые занимают особи, принадлежащие к разным половозрастным группам. При дифференциации ниш происходит снижение уровня конкуренции, в первую очередь, топической и трофической (таблица 7).
Таблица 7 – Примеры дифференциации внутривидовых ниш у животных
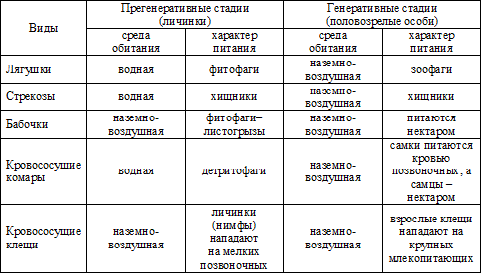
Генетическая структура – соотношение в популяции различных генотипов и аллелей. Совокупность генов всех особей популяции называют генофондом. Генофонд характеризуют частоты аллелей и генотипов. Частота аллеля – это его доля во всей совокупности аллелей данного гена. Сумма частот всех аллелей равна единице:
p + q = 1,
где p – доля доминантного аллеля (А), q – доля рецессивного аллеля (а).
Согласно закону Харди-Вайнберга, относительные частоты аллелей в популяции остаются неизменными из поколения в поколение. Закон Харди–Вайнберга справедлив, если соблюдаются следующие условия:
- популяция велика;
- в популяции осуществляется свободное скрещивание;
- отсутствует отбор;
- не возникает новых мутаций;
- нет миграции новых генотипов в популяцию или из популяции.
Очевидно, что популяций, удовлетворяющих этим условиям, в течение длительного периода времени, в природе не существует. На популяции всегда действуют внешние и внутренние факторы, нарушающие генетическое равновесие. Длительное и направленное изменение генотипического состава популяции, ее генофонда получило название элементарного явления. Без изменения генофонда популяции невозможен эволюционный процесс.
Генетическая структура популяции поддерживается степенью индивидуальной изменчивости особей. Чем генетически более разнородна популяция, тем выше ее экологическая пластичность. Считается, что поддержание стабильной генетической структуры обеспечивается случайным характером скрещивания, хотя во многих группах высших животных доминирующие (преобладающие) особи имеют преимущества в спаривании. Например, на лежбищах морских слонов (как и у многих других ластоногих) секачи-доминанты составляют всего 4 % от всех самцов, но принимают участие в 85 % спариваний и оплодотворяют до 85 % caмок.
Выделяют следующие факторы, изменяющие генетическую структуру популяции:
- мутации – источник возникновения новых аллелей;
- неравная жизнеспособность особей (особи подвергаются действию отбора);
- неслучайное скрещивание (например, при самооплодотворении частота гетерозигот постоянно падает);
- дрейф генов – изменение частоты аллелей случайные и независящие от действия отбора (например, вспышки заболеваний);
- миграции – отток имеющихся генов и (или) приток новых.
Основные характеристики популяций
Популяции обладают рядом свойств, которые не присущи отдельно взятой особи или просто группе особей. К основным характеристикам популяции относятся: численность, плотность, рождаемость, смертность, относительный и абсолютный прирост.
Численность – общее количество особей на данной территории или в данном объеме. Плотность – количество особей или их биомасса на единицу площади или объема. Сравнительно многочисленные популяции могут обладать низкой плотностью. Низкая плотность снижает внутривидовую конкуренцию и повышает шансы отдельных членов популяции на выживание. При низкой плотности популяция не оказывает заметного влияния на функционирование сообществ и экосистем; например, энергия, затраченная хищником на поиск жертвы, не компенсируется энергетическими выгодами от съеденной жертвы – тогда становится бессмысленной пищевая специализация хищника.
Возможна и обратная ситуация: сравнительно малочисленная популяция может обладать высокой плотностью. В этом случае гарантируется образование брачных пар. При малочисленности, но высокой плотности популяция может оказывать значительное влияние на функционирование сообществ и экосистем.
Таким образом, низкая плотность популяции уменьшает ее шансы на воспроизведение, но увеличивает шансы на выживание. Высокая плотность, наоборот, увеличивает шансы на воспроизведение, но уменьшает шансы на выживание. Следовательно, каждая конкретная популяция должна обладать некоторой оптимальной плотностью.
В природе происходит постоянные колебания численности и плотности популяции.
Динамика численности и плотности определяется в основном рождаемостью, смертностью и процессами миграции. Это показатели, характеризующие изменение популяции в течение определенного периода: месяца, сезона, года и т.д. Изучение этих процессов и причин их обусловливающих очень важно для прогнозов состояния популяций.
Рождаемость различают абсолютную и удельную. Абсолютная рождаемость – это количество новых особей, появившихся за единицу времени, а удельная – то же самое количество, но отнесенное к определенному числу особей. Относительная рождаемость может рассчитываться или на одну особь, или на 1000 особей. Например, в популяции в начале года было 10 000 особей, а в течение года родилось 156 новых особей; тогда удельная рождаемость равна 156/10000 = 0,0156 на одну особь, или 0,0156 * 1000 = 15,6 на тысячу особей.
Существуют моноциклические (у растений монокарпические) виды, представители которых размножаются один раз в жизни, и полициклические (у растений поликарпические) виды, представители которых размножаются неоднократно.
Рождаемость определяется многими факторами: условиями среды, наличием пищи, биологией вида (скорость полового созревания, количество генераций в течение сезона, соотношение самцов и самок в популяции). Так, рождаемость человека не очень высока в экономически развитых странах и примерно вдвое выше в развивающихся странах с недостаточным уровнем контроля за рождаемостью (таблица 8, рисунок 91).
Согласно правилу максимальной рождаемости (воспроизводства) в идеальных условиях в популяциях появляется максимально возможное количество новых особей; рождаемость ограничивается физиологическими особенностями вида. Например, одуванчик за 10 лет способен заполонить весь земной шар, при условии, что все его семена прорастут. Исключительно обильно семеносят ивы, тополя, березы, осина, большинство сорных растений. Бактерии делятся каждые 20 минут и в течение 36 часов могут сплошным слоем покрыть всю планету. Очень высока плодовитость у большинства видов насекомых и низка у хищников, крупных млекопитающих.
Экологическая рождаемость – дает представление о скорости возрастания численности популяции при фактически сложившихся условиях жизни рассматриваемой группы особей. Экологическая рождаемость не постоянна и изменяется в зависимости от физических условий среды и состава популяции.
В целом для видов, которые не заботятся о потомстве, характерна высокая максимальная и низкая экологическая рождаемость (взрослая самка трески выметывает миллион икринок, из которого в среднем доживают до взрослого состояния лишь 2 особи). У менее приспособленных к неблагоприятным условиям видов высокая смертность в молодом возрасте компенсируется значительной плодовитостью. В благоприятных условиях плодовитость обычно низкая. Этим объясняется более высокая плодовитость животных в зонах умеренного климата по сравнению с тропическими видами.
Характер плодовитости зависит: от скорости полового созревания, числа генераций в течение сезона, от соотношения в популяции самцов и самок. Если вид размножается с большой скоростью и чутко реагирует на изменения условий среды, то численность популяций его быстро и существенно меняется. Это относится ко многим насекомым и мышевидным грызунам.
Размеры популяции могут уменьшатся в результате эмиграции и смертности. Смертность, как и рождаемость, бывает абсолютной (количество особей, погибших за определенное время), так и удельной. Она характеризует скорость снижения численности популяции от гибели из– за болезней, старости, хищников, недостатка корма, и играет главную роль в динамике численности популяции.
Различают три типа смертности:
- одинаковая на всех стадиях развития; встречается редко, в оптимальных условиях;
- повышенная смертность в раннем возрасте; характерна для большинства видов растений и животных (у деревьев к возрасту зрелости доживает менее 1% всходов, у рыб – 1–2% мальков, у насекомых – менее 0,5% личинок);
- высокая смерть в старости; обычно наблюдается у животных, чьи личиночные стадии проходят в благоприятных мало изменяющихся условиях: почве, древесине, живых организмах.
Человеческая смертность самая низкая в развитых странах с высоким уровнем медицинского обслуживания (табл. 8, рис. 91).
Таблица 8 – Рождаемость и смертность человека в различных регионах

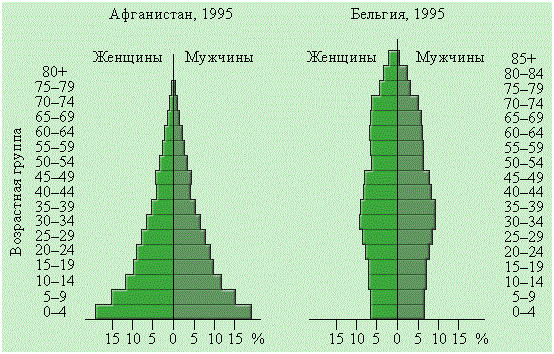
Рисунок 91 – Рождаемость и смертность в различных странах
Динамика популяций – это процессы изменений ее основных биологических показателей во времени.
В зависимости от соотношения рождаемости и смертности в популяции ее численность:
- либо растет;
- либо сохраняется на постоянном уровне;
- либо сокращается.
Ученым, исследующим динамику популяций, важно бывает знать не только общую смертность в популяции, но и количество особей, погибающих до достижения половой зрелости. Условием стабильности популяции является тот факт, что до половой зрелости доживают два потомка от каждой пары. Динамику смертности популяции в зависимости от возраста отображают на кривых выживания. Существуют три основных типа этих кривых: когда главным фактором смертности является естественное старение особей (на рисунке 92 эта кривая обозначена буквой А), когда в популяции высока ранняя смертность (Б) и, наконец, когда смертность постоянна в течение всей жизни организмов (В). Кривые выживания конкретных видов могут занимать промежуточные положения между этими крайними формами.
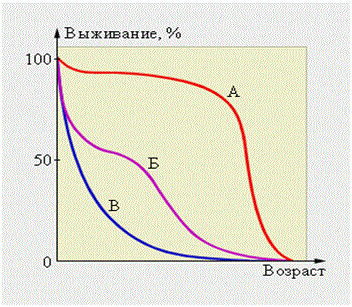
Рисунок 92 – Типы кривых выживания
Относительный прирост популяции представляет собой разность между относительной рождаемостью и относительной смертностью. Это важнейшая характеристика популяции (обычно обозначается символом r). Относительный прирост популяции может быть положительным, нулевым и отрицательным.
Для изолированной популяции: прирост = рождаемость – смертность.
Для открытой популяции: прирост = (рождаемость + иммиграция) – (смертность + эмиграция).
Абсолютный прирост популяции представляет собой изменение абсолютной численности популяции за бесконечно малый промежуток времени. Абсолютное изменение численности популяции в единицу времени обозначается выражением dN/dt.
Если рождаемость в изолированной популяции превышает смертность, то популяция растёт. В начале роста кривая роста популяции – это экспонента, описываемая J–образной кривой (рис. 93 А). Однако рано или поздно питательные запасы в окружающей среде исчерпываются. В одних популяциях уменьшение количества питательных веществ начинает воздействовать на динамику роста загодя, и кривая роста приобретает S–образную форму, происходит сигмоидный рост (рис. 93 Б). В популяциях экспоненциального типа бесконтрольный рост численности заходит слишком далеко, после чего происходит катастрофический «обвал» численности, связанный с истощением ресурсов. Рост популяций на первой (экспоненциальной) фазе можно приближенно описать дифференциальным уравнением
dN / dt = (b – d) N = rN,
где N(t) – количество особей в зависимости от времени, b – рождаемость, d – смертность, а r = b – d – врождённая скорость роста численности популяции.
Дифференциальным уравнением можно описать и S–образную кривую:
dN / dt = rN (K- N) / K,
где K – это максимальный размер стабильной популяции, которая может существовать неопределенно долго, если не изменятся внешние условия. Заметим, что дифференциальные уравнения роста – приблизительные уравнения, так как они не учитывают различия между особями, сезонные колебания количества пищи и способности к размножению и т. п.
Эти типы могут усложняться, модифицироваться или сочетаться (рис. 94).
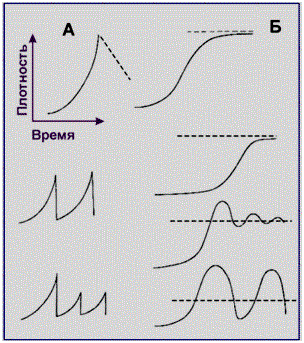
Рисунок 93 – Кривые роста численности популяции:
А – экспоненциальный рост; Б – сигмоидный рост
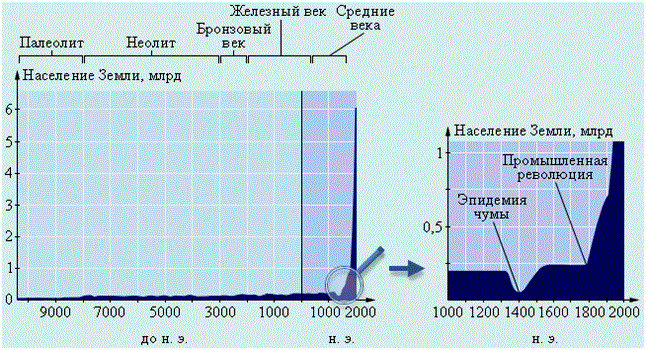
Рисунок 94 – Кривые численности популяции человека
Выделяют три типа динамики популяций: стабильный, изменчивый и взрывной (рис. 95). Стабильным считается ход численности при изменениях всего в несколько раз, изменчивым – при колебаниях в десятки раз, а взрывная динамика характеризуется периодическим превышением обычной численности в сотни и тысячи раз.
Рисунок 95 – Типы популяционной динамики:
А – стабильный; Б – изменчивый; В – взрывной
Например, циклы трех– и четырехлетней периодичности характерны для многих северных мышевидных грызунов (мышей, полевок, леммингов) и их хищников (полярной совы, песцов). В Европе лемминги порой достигают такой высокой плотности, что начинают мигрировать из своих перенаселенных местообитаний. Животные проходят через деревни в таком количестве, что кошки и собаки, которые вначале нападали на них, просто перестают обращать внимание на грызунов. Достигнув моря, многие лемминги тонут.
Типы популяционной динамики численности наиболее хорошо изучены у лесных насекомых, на примере которых выявляется большое многообразие конкретных связей и особенностей видов, влияющих на плотность их популяций.
В лесных сообществах преобладают виды со стабильным ходом популяционной динамики. Они питаются практически за счет всех частей деревьев (стволы, корни, побеги, листья, почки, плоды, семена), однако при этом не оказывают губительного воздействия на растения. Как правило, такие индифферентные виды отличаются низкой миграционной активностью, ведут оседлый образ жизни. Комплекс их паразитов включает обычно несколько сильно специализированных видов, эффективно поражающих популяцию хозяина.
Флюктуирующий тип динамики численности свойствен многим ксилофагам (потребителям коры и древесины): усачам, златкам, короедам. Для них характерно совместное заселение кормовых объектов – ослабление деревьев. Это позволяет быстро снизить устойчивость дерева, но в то же время совместное обитание ксилофагов обостряет конкурентные отношения между ними, что действует как безынерционный механизм регуляции численности.
Для короедов, поселяющихся первыми на ослабленных деревьях, при избыточном запасе питательных веществ (растворимых углеводов и крахмала) развитие лимитируется воздействием защитных реакций дерева, таких, например, как смоловыделение у хвойных. Кроме того, еще отсутствуют симбиотические микроорганизмы. Сопротивление еще жизнеспособных деревьев может быть сломлено при концентрированном нападении и избыточно высокой плотности вредителя. Другой предел пригодности дерева для короедов – полное отмирание и разрушение луба. Между двумя указанными состояниями дерева для короедов создается оптимум кормовых условий, отмечается максимальная выживаемость всех фаз развития и наиболее высокие коэффициенты размножения.
Относительную роль различных регуляторных механизмов, определяющих флюктуирующий ход численности лесных насекомых, можно проследить на примере большого лиственного короеда – Ips subelongatus. Этот вредитель лиственницы и частично других пород широко распространен по всей таежной зоне.
Жуки нападают на физиологически ослабленные живые лиственницы, у которых отсутствует активное защитное смоловыделение. Внедряясь под кору, самцы продуцируют запаховые вещества – феромоны, привлекающие других особей, чем обеспечивается массовость заселения подходящего дерева и его дальнейшее ослабление. Феромоны играют также роль химических регуляторов внутрипопуляционных отношений – распределения короедов по стволу, своего рода меткой территории отдельных гнезд. Жуки, не нашедшие места на данном дереве, переселяются на другие. В дальнейшем внутривидовая конкуренция регулируется самками, которые в зависимости от плотности поселения изменяют длину маточных ходов и количество откладываемых яиц. При возрастающем недостатке площади часть самцов и самок мигрирует на другие объекты. Личинка короеда за период развития потребляет 0,78 см6 луба. Если «кормовые площадки» личинок при высокой плотности поселения становятся меньше этого предела, среди них увеличивается смертность. Таким образом, внутривидовая конкуренция регулирует заселенность дерева на всех фазах жизни вредителя: на стадии взрослых жуков она ведет к усилению миграций, на стадии личинок – к гибели части популяции. Показатели размножения остаются высокими на всех этапах флюктуации численности популяции.
Естественных врагов данного вредителя можно разделить на пять групп, по особенностям их взаимодействия с короедом. - Враги, действующие в период лёта короедов, – птицы (синицы, дятлы и др.), ктыри, муравьи. Они уничтожают часть популяции, но одновременно и энтомофагов короеда, т. е. других его естественных врагов. Их регуляторная деятельность поэтому слабо эффективна.
- Хищные насекомые, уничтожающие короеда на всех фазах развития под корой (ряд пестрянок, чернотелок, карапузиков, хищных мух и клопов). При низкой и средней плотности вредителя хищники сдерживают рост его численности, при высокой же плотности короедов – способствуют повышению выживаемости их вследствие ослабления внутривидовой конкуренции.
- Паразиты, заражающие только личинок короедов, – бракониды, хальциды. На регуляцию численности воздействуют слабо, так как пораженные личинки погибают лишь в конце жизни и общая смертность их от паразитов составляет несколько процентов.
- Паразиты, заражающие взрослых жуков, – нематоды, некоторые перепончатокрылые. Их влияние проявляется в снижении плодовитости и сокращении продолжительности жизни жуков, регулирующий эффект в целом невысок.
- Бактериальные и грибные заболевания. Эпизоотии среди короедов возникают лишь при резком ухудшении условий, когда действуют более мощные механизмы снижения численности.
Таким образом, в межвидовых отношениях короедов более или менее значительную роль в регуляции их численности имеют лишь хищные насекомые при относительно невысокой численности вредителя.
Способность к вспышкам массового размножения свойственна относительно немногим видам лесных насекомых. В сибирской тайге у скрытоживущих видов взрывной тип динамики численности характерен для алтайского лиственничного усача, большого черного хвойного усача, лиственничной почковой галлицы и некоторых других. Среди открытоживущих листогрызущих насекомых способность давать вспышки массового размножения свойственна лишь некоторым видам чешуекрылых и перепончатокрылых (пилильщикам, ткачам). Отличительные черты экологии таких видов: высокая выживаемость в сильно изменчивой среде благодаря специальным адаптациям, высокой миграционной активности, высокой и изменчивой плодовитости. У открытоживущих видов часто выявляется эффект группы и фазовая изменчивость.
Один из опаснейших вредителей хвойных пород – сибирский шелкопряд Dendrolimus sibiricus, распространенный от Урала до Тихого океана. Этот вид обладает следующими внутрипопуляционными механизмами регуляции численности.
В очагах сибирского шелкопряда по фазам вспышки существенно изменяется соотношение полов. Доля самок варьирует от 32 до 76 %. При нарастании вспышки доминируют самки, при затухании – самцы. В переуплотненных популяциях повышается смертность самок на всех фазах развития, а также отмечается их более высокая миграционная активность из очагов размножения. В фазе максимума численности на периферии очага доля самок составляет до 73 %, а в центре – 44 %.
У сибирского шелкопряда, как и у других видов с вспышками массового размножения, плодовитость изменяется в широких пределах. Этот показатель сильно зависит от плотности популяции (рис. 96). При низкой и сверхвысокой численности плодовитость самок снижается почти втрое. Наиболее высокая плодовитость наблюдается в фазе нарастания численности (свыше 300 яиц на самку), а затем закономерно снижается под влиянием конкуренции и недостатка корма в переуплотненных очагах. Изменение соотношения полов и плодовитости можно рассматривать как безынерционные или слабоинерционные механизмы регуляции численности. Их действие проявляется в течение жизни одного поколения.
Рисунок 96 – Фазовый портрет динамики численности сибирского шелкопряда в Красноярском крае (из А. С. Исаева и др., 2001):
по вертикали – логарифм коэффициента размножения;
по горизонтали – логарифм плотности популяции гусениц на одно дерево. Схема иллюстрирует цикличность вспышек и зависимость показателей размножения от плотности популяции
Миграционная активность сибирского шелкопряда – также эффективный внутрипопуляционный механизм регуляции численности. У этого вида отмечены разные типы массовых перемещений. В период низкой численности пульсирующие миграции перераспределяют вредителя на площади лесных массивов, в результате чего формируются резервации – места сохранения вида. В основном это перестойные пихтовые леса на плакорах или склонах южной экспозиции. В начальной фазе вспышки происходит стягивание вредителя с больших территорий на ограниченный участок леса с наиболее благоприятными условиями (конденсирующие миграции). К концу фазы нарастания численности начинается вылет бабочек из первичных очагов, где к этому времени плотность популяции достигает критических величин. На фазе максимума численности происходят массовые выселения бабочек и их перемещение воздушными потоками на расстояния до 100 км. Среди мигрантов преобладают самки с запасом зрелых яиц. В редких случаях отмечаются активные массовые миграции бабочек. При затухании очагов сибирского шелкопряда развиваются дисперсионные миграции – рассеивание популяции из очагов размножения в окружающие лесные массивы.
Для сибирского шелкопряда известен также эффект группы – положительное влияние оптимальной плотности, которое проявляется в основном у гусениц I–III возрастов и выражается в ускорении их развития более чем в 2 раза. В регуляцию хода численности вредителя вносит вклад также гетероцикличность – наличие особей с одно– и двухлетним циклом в зависимости от плотности популяции.
Повышение плотности очага сильно ускоряет метаболические процессы у членов популяции, что выражается как в ускорении сроков развития, так и в изменении поведения и внешнего вида бабочек и гусениц – появляются особи с темной окраской. В разных фазах цикла массового размножения количество темных бабочек в лесах Средней Сибири изменяется от 10 до 80 %.
Среди естественных врагов сибирского шелкопряда отмечено более 60 видов. Однако, несмотря на разнообразие, энтомофаги не могут постоянно держать под контролем численность вредителя из–за высокой инерционности их количественной реакции. Усложняющий фактор – высокая активность сверхпаразитов. Многие энтомофаги сибирского шелкопряда гораздо менее его устойчивы к факторам абиотической среды. Например, один из эффективных паразитов яйцеед–теленомус в массе гибнет в холодные зимы. Вот почему вспышки размножения сибирского шелкопряда часто возникают после холодных малоснежных зим.
При низкой численности сибирского шелкопряда в резервациях за счет естественных врагов–энтомофагов гибнет до 95 % яиц, 40 % гусениц и 70 % куколок. Основными регуляторами при этом являются яйцеед–теленомус и тахина-мазицера. Общие показатели смертности в популяциях достигают более 99 %. Это свидетельствует о высоких возможностях вредителя переходить к вспышке. Быстрый рост численности может быть обеспечен даже небольшим (5–10 %) увеличением выживаемости. На фазе нарастания численности поражаемость от энтомофагов падает (яйца – до 50 %, гусеницы – до 11 %, куколки – до 23 %). Энтомофаги «не успевают » сдерживать растущую популяцию вредителя. В период кульминации вспышки пресс энтомофагов на популяцию шелкопряда усиливается и достигает наибольшего выражения в период разреживания и депрессии численности, когда естественные враги полностью уничтожают избыток особей.
Таким образом, при массовых вспышках размножения лесные насекомые-вредители выходят из–под межвидового контроля и их возврат к стабильной численности происходит в основном в результате внутрипопуляционных регулирующих механизмов, которые наиболее эффективно действуют лишь при очень высоких уровнях плотности, губительной для лесного насаждения.
Понимание и учет всех этих факторов, определяющих динамику численности лесных вредителей, совершенно необходимы для принятия действенной системы мер борьбы с ними и профилактики вспышек, для определения необходимости, сроков, доз и форм применения средств, сдерживающих массовое размножение. С экологической точки зрения задача заключается в удержании популяции в зоне депрессии численности.
Понимание механизмов, определяющих движение численности популяций, дает в руки ключ управления ими. Однако эта задача не может решаться трафаретно для разных видов. Во всех случаях необходимо детальное знание экологических особенностей конкретного вида и условий его биотического окружения, а также характер взаимодействия модифицирующих факторов среды.
Все факторы, оказывающие влияние на численность популяции разделяют на две группы:
- не зависящие от плотности, если их влияние не зависит от величины популяции;
- зависящие от плотности, если их влияние на популяцию есть функция плотности.
Вторую группу факторов часто называют регуляторными, или управляющими плотностью. Действие абиотических факторов часто не зависит от плотности популяции, тогда как действие биотических факторов (конкуренция, хищничество, паразитизм) часто зависит от плотности.
При благоприятных условиях в популяциях наблюдается рост численности и может быть столь стремительным, что приводит к популяционному взрыву. Совокупность всех факторов способствующих росту численности называется биотическим потенциалом. Этот показатель отражает теоретический максимум потомков от одной пары (или одной особи) за единицу времени, например год или весь жизненный цикл.
Величина биотического потенциала чрезвычайно различна у разных видов. Напимер, самка косули способна произвести за жизнь 10–15 козлят, самка медоносной пчелы – 50 тыс. яиц, рыба–луна – до 3 млрд. икринок. Биотический потенциал этих видов на самом деле еще больше, так как часть развивающихся яиц и зародышей погибает еще до рождения. Если бы все зародыши сохранялись, а все потомство выживало, численность любой популяции через определенные интервалы увеличивалась бы в геометрической прогрессии.
Совокупность факторов, лимитирующих рост численности популяции, называют сопротивлением среды. Сопротивление среды сильнее всего действует на молодых особей, больше других страдающих от нападения хищников, болезней, недостатка воды и пищи или других неблагоприятных условий.
Состояние равновесия между биотическим потенциалом вида и сопротивлением среды (рис. 97), поддерживающее постоянство численности популяции получило название гомеостаза или динамического равновесия. Понятие гомеостаза является одним из важнейших в экологии. По отношению к любой природной системе (популяция, биоценоз, экосистема) гомеостаз определяется как состояние ее внутреннего функционального равновесия, поддерживаемое постоянным саморегулированием компонентов. Гомеостаз является характерным и необходимым для всех природных систем – от атома и организма до космических образований. При нарушении его происходят колебания численности популяции.
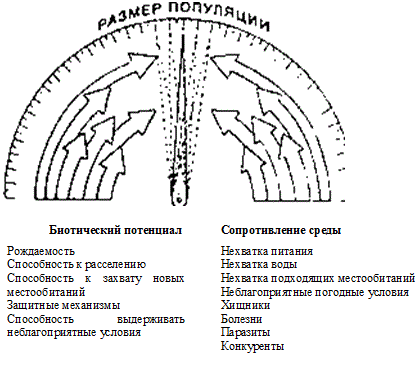
Рисунок 97 – Равновесное состояние популяции (по Б.Небелу, 1993)
Влияние ослабления или усиления пресса хищников на динамику популяций и структуру сообществ. Ярким примером того, к чему приводит снятие пресса потребителей на популяции жертв, являются масштабные «нашествия» чужеродных видов, попадающих в подходящие для размножения условия на других материках. На родине такие виды испытывают влияние активных регуляторов, занимают соответственные экологические ниши в биоценозах и, как правило, не считаются вредителями в хозяйственной практике человека. В новой среде, не имея специализированных врагов, эти виды за короткие сроки дают взрывы численности, нарушая структуру сложившихся сообществ. С экологических позиций впервые обзор и анализ таких событий был сделан английским зоологом Ч. Элтоном, одним из основоположников популяционной экологии. Вводя понятие «взрыв» применительно к непомерному увеличению численности какого–либо вида, Ч. Элтон объяснял, что он намеренно употребляет это слово, так как оно обозначает «внезапное освобождение сил, проявление которых сдерживалось ранее другими силами».
«Нашествия» иноземных видов, занос которых преднамеренно или случайно осуществляется человеком, широко распространены в современном мире и часто вызывают большие осложнения в хозяйственной деятельности. Относительно немногие примеры успешной борьбы с ними связаны, как правило, с интродукцией естественных врагов, т. е. воссозданием системы биоценотической регуляции. Если на «захваченной» видом территории постепенно складывается новый комплекс регуляторов, тогда его численность идет на убыль. Однако для некоторых вселенцев этот процесс происходит достаточно медленно. Таким примером является опасный вредитель картофеля колорадский жук Leptinotarsa septemlineata, распространение которого в Европе началось после заноса из Америки в 20–х годах прошлого века с западных берегов Франции. Постепенно распространяясь на восток, колорадский жук занял в настоящее время всю европейскую территорию, картофелеводческие районы Сибири, обнаружен на Дальнем Востоке и острове Сахалин (рис. 98). Жук и его личинки имеют ядовитую гемолимфу, спасающую их от насекомоядных птиц и других позвоночных. У колорадского жука на евроазиатской территории до сих пор не сформировался эффективный комплекс потребителей. Потери урожая картофеля от колорадского жука составляют в среднем до 40 %.
Рисунок 98 – Распространение колорадского жука по территории России (на 1990 г.)
Другим примером современного «нашествия» вида, изменившего всю структуру сообщества, является размножение в Черном море гребневика Mnemiopsis leigyi, занесенного с балластными водами судов от побережья Северной Америки (рис. 99). Этот гребневик обладает гигантским репродуктивным потенциалом, производя ежесуточно до 10 000 яиц. Личинки тоже могут продуцировать яйца, из которых развиваются взрослые гребневики. При наличии пищи общая масса мнемиопсиса может удваиваться за 2–3 суток. Гребневик питается ветвистоусыми и веслоногими рачками, другим мелким зоопланктоном, икрой и личинками рыб. Впервые мнемиопсис был отмечен в Черном море в 1987 г. Уже через три года его масса составила 3–4 кг, а местами до 12 кг под каждым квадратным метром воды. Одновременно резко упала численность медуз Aurelia aurita, господствовавших до этого в планктоне (рис. 100). Выедая кормовой планктон, икру и личинок рыб, гребневик вызвал резкое падение общего их улова. Например, вылов ставриды сократился, со 110–115 тыс. т всего до 3 тыс. т. В Азовском море запасы тюльки – корма для хищных рыб – сократились в 400 раз, а ее добыча упала с 50–70 тыс. т до 100 т. В последующие годы численность гребневика понизилась, но он по-прежнему выедал основную массу зоопланктона (рис.101). С 1997 г. в Черном море стал встречаться другой гребневик – Веrое ovata, распространенный на севере Атлантики и в Баренцевом море. Для него мнемиопсис явился хорошей кормовой базой, и численность последнего упала на порядок величин. В некоторых местах акватории он вообще не встречался. Снова стала размножаться медуза аурелия. Однако обилие Beroe в Черном море неустойчиво, этот гребневик адаптирован к более высокой солености воды. Появление и размножение его происходит волнами, а в период спада снова размножается Mnemiopsis.
Рисунок 99 – Гребневик мнемиопсис – внешний вид (из И. Е. Виноградова и Э. А. Шукшиной, 1993)
Рисунок 100 – Колебания биомассы аурелии и мнемиопсиса в глубоководной части Черного моря (в пересчете на углерод, по И. Е. Виноградову и Э. А. Шукшиной, 1993)
Рисунок 101 – Колебание биомассы в открытых районах Черного моря некоторых групп планктонных организмов, которыми питается мнемиопсис (по И. Е. Виноградову и Э. А. Шукшиной, 1993): - саггиты;
- веслоногий рачок Calanus luxinus;
- мелкие рачки и личинки донных животных
Чрезмерное усиление пресса хищников на популяцию какого–либо вида в сообществе может отозваться через сеть трофических зависимостей на довольно далеких уровнях. Например, можно выстроить цепь причинных связей между усилением промысла минтая и других рыб в районе Берингова моря и исчезновением зарослей прибрежных водорослей – ламинарий. Усиленный рыбный промысел (при котором человек выступает в роли мощного хищника) стал одной из причин падения численности питающихся рыбой сивучей и нерпы – основной добычи крупных хищных дельфинов – косаток. С 1991 г. косатки стали охотиться в прибрежных водах на каланов, на которых не нападали раньше, добывая пищу в открытом океане. С начала 90-х годов на Алеутских островах исчезло 40 тыс. каланов, являющихся заповедным видом. Численность этих зверей за 7 лет сократилась почти в 10 раз. Каланы питаются в основном морскими ежами, обилие которых в результате возросло тоже на порядок величин. Морские ежи – главные враги бурых водорослей, заросли которых у побережья сократились за 10 лет в 12 раз.
В природе численность популяций испытывает колебания. В связи с размерами ареала популяций может значительно изменяться и численность особей в популяциях. Так, у насекомых и мелких растений открытых пространств численность особей в отдельных популяциях может достигать сотен тысяч и миллионов особей. Напротив, популяции животных и растений могут быть и сравнительно небольшие по численности.
Различают периодические и непериодические колебания численности популяций. Первые совершаются в течение сезона или нескольких лет (4 года – периодический цикл плодоношения кедра, подъема численности лемминга, песца, полярной совы; через год плодоносят яблони на садовых участках) (рис. 102), вторые – это вспышки массового размножения некоторых вредителей полезных растений, при нарушениях условий среды обитания (засухи, необычно холодные или теплые зимы, слишком дождливые сезоны вегетации), непредвиденные миграции в новые местообитания (рис. 103). Периодические и непериодические колебания численности популяций под влиянием биотических и абиотических факторов среды, свойственные всем популяциям, именуются популяционными волнами.
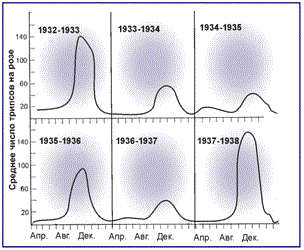
Рисунок 102 – Сезонные изменения численности популяции взрослых трипсов на розах (Davidson, Andrewartha, 1948)
Рисунок 103 – Рост дрожжей в культуре (Allee et al., 1949)
Популяции многих видов организмов способны к саморегуляции своей численности. Выделяют три механизма торможения роста численности популяций:
- при возрастании плотности повышается частота контактов между особями, что вызывает у них стрессовое состояние, уменьшающее рождаемость и повышающее смертность;
- при возрастании плотности усиливается эмиграция в новые местообитания, краевые зоны, где условия менее благоприятны и смертность увеличивается;
- при возрастании плотности происходят изменения генетического состава популяции, например, быстро размножающиеся особи заменяются медленно размножающимися.
Понимание механизмов регуляции численности популяций чрезвычайно важно для возможности управления этими процессами. Деятельность человека часто сопровождается сокращением численности популяций многих видов. Причины этого в чрезмерном истреблении особей, ухудшении условий жизни вследствие загрязнения окружающей среды, беспокойства животных, особенно в период размножения, сокращение ареала и т.д. В природе нет и не может быть «хороших» и «плохих» видов, все они необходимы для ее нормального развития. В настоящее время остро стоит вопрос сохранения биологического разнообразия. Сокращение генофонда живой природы может привести к трагическим последствиям. Международный союз охраны природы и природных ресурсов (МСОП) издает «Красную книгу», где регистрирует следующие виды: исчезающие, редкие, сокращающиеся, неопределенные и «черный список» безвозвратно исчезнувших видов.
В целях сохранения видов человек использует различные способы регулирования численности популяции: правильное ведение охотничьего хозяйства и промыслов (установление сроков и угодий охоты и отлова рыбы), запрещение охоты на некоторые виды животных, регулирование вырубки леса и др.
В то же время деятельность человека создает условия для появления новых форм организмов или развития старых видов, к сожалению, часто вредных для человека: болезнетворных микроорганизмов, вредителей сельскохозяйственных культур и т.д.
При всем многообразии механизмов популяционного гомеостаза его функции можно сгруппировать в три категории (Шилов, 1997):
- поддержание пространственной структуры;
- поддержание генетической структуры;
- регуляция численности и плотности населения.
Механизмы поддержания пространственной структуры разнообразны. Наиболее важные из них следующие.
Территориальная агрессия – наиболее прямая форма территориальных взаимоотношений. Животные, кормящиеся на своем участке, проявляют агрессию при защите его от соседей. Птицы активно защищают территорию своего гнездования, грызуны охраняют друг от друга места своего пропитания, хищники заботятся об охране своих охотничьих участков.
Маркирование территории – относительно «мягкий» механизм гомеостаза, не связанный с агрессией. У птиц это выражается в акустической маркировке участков. Некоторые обитатели водной среды также прибегают к звуковым сигнализациям. Так, голубые киты при общении друг с другом издают низкочастотные импульсы порядка 188 дБ, что позволяет считать подобные звуки самыми громкими звуками, которые способны издавать животные. Их можно услышать на расстоянии 860 км.
У рыб нередко окраска тела говорит чужаку о занятости территории. Разнообразны механизмы маркировки территории у млекопитающих. Многие животные для мечения участков используют запах мочи и экскрементов. Но наибольшее значение имеют выделения особых желез. Например, африканской гориллы, полевки, песчанки и другие виды мышевидных грызунов.
Нередко в качестве меток используются визуально различимые знаки. Например, следы когтей бурого медведя на стволах деревьев, либо остатки шерсти, которыми многие представители семейства волчьих метят свою территорию.
Нежелательны любые отклонения параметров популяций от оптимальных, но если чрезмерно высокие значения их не представляют прямой опасности для существования вида, то снижение до минимального уровня, особенно численности популяции, представляют угрозу для вида.
Принцип минимального размера популяций: популяция не может состоять из меньшего числа особей, чем необходимо для ее стабильного развития и устойчивости к факторам внешней среды. Минимальными размерами популяций характеризуются очень многие виды на Дальнем Востоке: тигр амурский, леопард дальневосточный, белый медведь, утка-мандаринка, многие бабочки: хвостоносец Мака и хвостоносец Ксута, адмирал, зефиры, красавица Артемида, Аполлон, реликтовый усач, жук-олень; из растений: все аралиевые, орхидные, пихта цельнолистная, сосна густоцветковая, абрикос маньчжурский, можжевельник твердый, тис остроконечный, лилии двурядная, мозолистая, даурская и др., рябчик уссурийский, триллиум камчатский и многие другие виды.
Однако наряду с принципом минимального размера популяций есть и принцип, или правило, популяционного максимума (Ю.Одум, 1975). Популяции эволюционируют так, что регуляция их плотности осуществляется на значительно более низком уровне по сравнению с верхним пределом емкости местообитания, достигаемым лишь в том случае, если полностью используются ресурсы энергии и пространства. При росте плотности популяции снижается обеспеченность пищей. У многих животных от потребления пищи прямо зависит плодовитость: при увеличении плотности популяции плодовитость падает, и это предотвращает дальнейший рост численности. Лишь теоретически популяция способна к неограниченному росту численности.
Правило популяционного максимума конкретизируют два обобщения. Первое из них известно как теория лимитов популяционной численности, или теория X. Г. Андреварты – Л. К. Бирча (1954): численность естественных популяций ограничена истощением пищевых ресурсов и условий размножения, недоступностью этих ресурсов и слишком коротким периодом ускорения роста популяции. Второе обобщение дополняет первое. Оно носит название теории биоценотической регуляции численности популяции К. Фридерихса (1927): регуляция численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида. Совершенно очевидно – теория К. Фридерихса объясняет, что лимитирует и формирует потенцию роста численности популяции, заложенную генетически. Однако, как известно, бывают взрывы роста численности популяций – сбои в регуляторных системах самоограничения и внешних ограничений популяционного роста.
Несколько уже, чем у Ю. Одума и в упомянутых теориях, подход к взаимосвязи популяции и среды ее обитания у В. Уини–Эдвардса (в ряде случаев пишут Вини–Эдвардс), который сформулировал правило пищевой корреляции: в ходе эволюции сохраняются только те популяции, скорость размножения которых скоррелирована с количеством пищевых ресурсов среды их обитания. Важно подчеркнуть, что скорость размножения в норме всегда ниже максимально возможной, и постоянно остается запас пищевых ресурсов. Очевидно, что размер видового населения в ходе эволюции должен быть ограничен не текущими пищевыми возможностями, а их минимумом, возникающим или могущим возникнуть в течение достаточно большого ряда лет. В противном случае популяция столкнется с резким недостатком пищи и в значительной своей части вымрет как под воздействием прямого недостатка кормовых объектов, так и в результате межвидовой конкуренции, осложняемой биоценотическими регулирующими механизмами в целом (к этому вопросу мы еще вернемся в конце параграфа при обсуждении принципа стабилизации экологических ниш). Для того чтобы избежать подобной коллизии, и была эволюционно выработана преадаптация более низкой плотности населения популяции, чем допускают наличные кормовые возможности.
Конечно, при обсуждении проблем саморегуляции ограничивать действие регулирующих факторов среды одними лишь условиями питания нельзя. Следует говорить о емкости среды в целом, как это сделал Ю. Одум. Однако кормовые возможности играют одну из ведущих ролей и прежде всего бросаются в глаза. Их легче рассчитать математически.
Несколько шире ставит вопрос Д. Лэк, сформулировавший в 1954 г. принцип, или теорию, носящую его имя, теория, или принцип Лэка: среднее количество яиц, откладываемых птицами, эволюционно ограничено возможностями выращивания выведшихся из них птенцов. Очевидно, этот лимит связан с комплексом условий: интенсивностью обмена веществ, длиной дня, наличием пищи, ее близостью и т. д. В основе, безусловно, лежит энергетика системы – она отработана эволюцией вида в конкретных условиях его обитания. Энергетическая расточительность в природе слишком большая роскошь, хотя бывают и исключения.
Хорошо известно, что в ряде случаев происходят массовые размножения организмов, не предотвращаемые ни внутрипопуляционными, ни межвидовыми механизмами. При этом не срабатывает система эволюционной взаимосвязи: плотность населения животных как фактор отбора для растений – изменение генетической конституции растений, а также обратная связь: генетическая конституция растений – плотность населения животных. К тому же популяционные механизмы саморегуляции в мире животных никак нельзя переносить на популяции растений. У последних нет ни территориального, ни группового поведения, аналогичного тому, что существует у животных. При переходе к человеческому обществу приходится констатировать, что биологическая превентивная саморегуляция численности у него, как правило, отсутствует. Иначе бы в истории людей не возникли локальные, региональные и глобальные экологические кризисы. Очевидно, мы имеем дело с некой эволюционной нормой саморегуляции и закономерными отклонениями от нее.
В растительном мире, как правило, наблюдается максимальное заполнение пространства и использование солнечной энергии при сохранении запасов минерального питания. Лимитирующими факторами служат физическая заполненность территории, поток лучевой энергии, предшествующие события, материализованные в процессах почвообразования, собственное воздействие растений на среду (например, аллелопатия) и сложные взаимосвязи с представителями других царств природы – микроорганизмами, грибами и животными. Превентивных механизмов саморегуляции плотности населения не наблюдается. Однако существует мощный рычаг ее восстановления в случае изреживания – почвенный запас семян, спящие почки и т. п. И параллельно этому имеется генетический механизм ограничения размеров индивидов. В противном случае одно растение было бы способно занять всю площадь и не оставалось бы места для процессов дублирования как инструмента обеспечения надежности системы. Емкость местообитания используется растениями максимально полно в рамках совокупности экологических ограничений. Основной их биотический механизм перенесен в область взаимодействия собственных размеров, межвидовых отношений и индивидуального воздействия на среду жизни. Свободный запас вещества и энергии минимален. В основном действуют факторы, непосредственно зависящие от плотности населения популяции.
Однако по отношению к минеральному питанию, т. е. к «пище» растений, они ведут себя подобно животным, имея механизм эволюционной преадаптации, препятствующий истощению почв. Этот механизм ориентирован на скорость почвообразовательных и шире — биогеоцентрических процессов. В одних случаях запас велик (например, черноземы), в других он очень мал (тропические леса). Чем интенсивнее круговорот веществ в экосистеме, тем ниже положительный баланс соотношения ресурсов минерального питания растений и скорости роста их популяций. Растения «берут в основном числом, а не умением».
В царствах дробянок, грибов и животных, как правило, существует иная закономерность: потенциальный запас жизненных ресурсов огромен – обычно используется не более 30 – 60 % наземной луговой растительности (крупные млекопитающие) и 10 % лесной растительности. Более интенсивно и полно утилизируется растительноядными животными чистая продукция водных растений. Как хорошо известно, пирамида биомасс в водной среде как бы перевернута – продуценты размножаются столь интенсивно, что консументы могут превышать их по биомассе. Однако это касается лишь мелких обитателей водной среды. Для рыб, птиц и млекопитающих общая их биомасса всегда намного ниже, чем потребляемых ими растительных кормов.
Случаи массового размножения организмов следует отнести к двум группам. Одна из них связана с антропогенными трансформациями среды жизни. Например, в сибирской тайге очаги массового размножения вредителей никогда не возникают в коренных лесах. Другая группа, видимо, связана с особым типом отношений между популяционной динамикой и экосистемными механизмами. Эволюционный отбор идет не на уровне внутрисистемных отношений в рамках собственной экосистемы, а на межэкосистемном уровне. Таковы вылеты саранчи, некоторых бабочек, отчасти миграции леммингов, белок, из птиц – кедровок.
В первом случае массовое размножение стимулируется аномальными для популяции условиями жизни: повышением потока энергии сверх обычного уровня, необычными условиями питания, снижением пресса конкурентов, хищников и паразитов, особо благоприятными условиями гидрологического режима и т. п. Переразмножение в изначальном очаге перебрасывается на широкие площади и затем резко обрывается с гибелью значительной части популяции от недостатка кормов или с развитием болезней и пресса других врагов.
Во втором случае арена действия принципиально схожа, но значительно пространственно шире. Аналогия в том, что вспышка массового размножения возникает в одних экосистемах и перекидывается на другие. Но имеется существенная разница. Вместо узких очагов переразмножения – обширное его поле. Вместо локального воздействия – широко межрегиональное, значительные выселения особей (пешая саранча, переселения, часто безвозвратные, млекопитающих и птиц). Как и в первом случае, затухание происходит как в изначальном ареале массового размножения, так и вне его, но в первом случае в сходных с очагом размножения экосистемах, а во втором часто – в резко отличных. И в этом втором случае действуют более высокие по иерархии экосистемы, «тушащие популяционный пожар».
Здесь уместно упомянуть одно частное правило, а именно правило стрессогенного увеличения надпочечников у высших позвоночных животных Я. Кристиана. Заключается оно в том, что в условиях перенаселенности у высших позвоночных животных увеличиваются надпочечники. Это результат сдвига нейроэндокринного гомеостаза, ведущего к изменению поведения животных (в частности, к повышению их агрессивности), репродуктивного потенциала (происходит резорбция эмбрионов), снижению устойчивости к заболеваниям и другим последствиям. Комплекс этих изменений обычно ведет к резкому падению жизнеспособности особей, их массовой гибели, снижению плотности населения популяции или даже ее элиминации от «стрессового шока». Как ясно из сказанного, основой обсуждаемого правила служит теория общего адаптационного синдрома Г. Селье.
Частное наблюдение стрессогенного увеличения надпочечников вкупе с теорией общего адаптационного синдрома Г. Селье вылилось в теорию Дж. Кристиана – Д. Дейвиса, которые в работах 1950–1968 гг. объяснили регуляцию численности популяций млекопитающих их социальным поведением, отражающимся на эндокринных реакциях (что и ведет к увеличению надпочечников).
Позвоночные, да и беспозвоночные животные в норме не могут абсолютно разрушительно воздействовать на среду своего обитания (правило сохранения видовой среды обитания), поскольку такое разрушение лишает их собственной основы жизни. Грубо говоря, съесть сразу все означает затем остаться без пищи и погибнуть от голода. Естественно, эволюционно должны были возникнуть ограничивающие механизмы популяционного и внепопуляционного характера. С одной стороны они обеспечивают возможность поиска (или поиска для неживотных) корма, с другой – ограничивают такой поиск или снижают саму плотность населения через популяционные механизмы – физико–химические и поведенческие. Для нас сейчас не столько важны эти механизмы, как сама констатация того, что в нормальных условиях действует закон (правило) популяционного максимума и правило пищевой корреляции, имеющее множество модификаций в зависимости от среды жизни (водной, наземной и т. д.) и от царства природы, даже систематического типа и класса животных (членистоногие, млекопитающие и т. д.).
Группа закономерностей саморегуляции в системах живого имеет важное практическое и теоретическое значение. Виды (популяции) поддерживают собственную среду жизни, то же происходит в рамках биоценозов и других биотических образований, в том числе на уровне всего живого вещества планеты. Если механизмы самоподдержания на популяционном уровне относительно независимы от деятельности человека, то деформируя биоценозы, человек сильно видоизменяет гомеостаз на уровне ценозов. Разрушение этого механизма в биосфере означало бы глубокую трансформацию жизни или даже ее гибель.
Огромная экологическая литература подтверждает правило колебаний (цикличности) численности: никакая популяция не находится в состоянии абсолютной уравновешенности числа особей, обязательно помимо сезонных изменений численности особей возникают периодические флуктуации, обусловленные внешними по отношению к популяции факторами, и осциляции, связанные с собственными (внутренними) динамическими изменениями популяции. Совершенно очевидно, что раз имеются ограничения согласно действию принципа минимального размера популяций и закона популяционного максимума, должно существовать обобщающее правило максимума размера колебаний плотности популяционного населения. Ю. Одум так формулирует это правило: «Существуют определенные верхние и нижние пределы для размеров популяции, которые соблюдаются в природе или которые теоретически могли бы существовать в течение сколь угодно длительного отрезка времени». В условиях стабильности среды обитания может быть 100 птиц на 1 га и 20 000 почвенных членистоногих на 1 м2, но никогда не бывает 20 000 птиц на 1 м2 и всего 100 членистоногих на 1 га. Правило максимума размера колебаний плотности популяционного населения может быть названо также законом количественной константности популяционного населения.
Кроме экологических факторов, включаются внутренние (генетические и физиологические) механизмы регулирования численности популяций: при сокращении жизненного пространства и запасов корма сокращается плодовитость особей (многие насекомые, мышевидные грызуны), повышение смертности на ранних стадиях жизни (паразиты, многие насекомые), задерживается наступление половой зрелости (полевые мыши) и стадии плодоношения (виды деревьев 2 и 3 ярусов в густом лесу), имеет место каннибализм (грызуны, насекомые, рыбы), и др.; снижается выход личинок из яиц (майский хрущак), уменьшаются размеры взрослых особей. При чрезмерном росте численности популяции у млекопитающих, общественных насекомых, птиц начинается эмиграция на новые места.
Экологические стратегии популяций
Приспособления организмов, в конечном итоге, направлены на повышение вероятности выжить и оставить потомство.
Экологическая стратегия популяции – это ее общая характеристика роста и размножения. Экологическая стратегия зависит также и от особенностей вида, и от условий среды. Экологические стратегии очень разнообразны. Однако среди этого многообразия можно выделить два крайних типа, которые называются r– стратегией и К–стратегией (рис. 115).
r-стратегия – определяется отбором, направленным прежде всего на повышение скорости роста популяции и, следовательно, таких качеств, как высокая плодовитость, ранняя половозрелость, короткий жизненный цикл, способность быстро распространяться на новые местообитания и пережить неблагоприятное время в покоящейся стадии.
К–стратегия – направлена на повышение выживаемости в условиях уже стабилизировавшейся численности. Это отбор на конкурентоспособность, повышение защищенности от хищников и паразитов, повышение вероятности выживания каждого потомка.
r–Стратеги (эксплеренты) характеризуются низкой конкурентоспособностью, высокой плодовитостью, отсутствием заботы о потомстве, быстрым развитием и короткой продолжительностью жизни. r–Стратегов образно называют «шакалами», поскольку они способны за короткое время завоевывать освободившееся экологическое пространство.
Популяции видов, у которых рождаемость и смертность в значительной мере зависят от их плотности (то есть от характеристики самой популяции), в меньшей степени зависят от действия внешних факторов. Они поддерживают численность, близкую к величине К, поэтому способ воспроизведения таких популяций называется К–стратегия (таблица 3).
К–Стратеги (виоленты) характеризуются высокой конкурентоспособностью, низкой плодовитостью, заботой о потомстве, длительным развитием и длительной продолжительностью жизни. К–Стратегов образно называют «львами», поскольку они способны долгое время удерживать экологическое пространство.
Рисунок 104 – K и r– стратегии популяций
Кроме r–стратегии и К–стратегии выделяется еще и S–стратегия. S–стратеги (патиенты) населяют местообитания с неблагоприятными условиями жизни для большинства организмов, в которых конкуренция практически отсутствует. Поэтому S–стратегов образно называют «верблюдами». По низкому значению r они близки к «львам» (виолентам), а по высокому значению К – к «шакалам» (эксплерентам). По длительности развития и длительности жизни S–стратеги могут быть сходными и с r–стратегами, и с К–стратегами.
Очевидно, что каждый организм испытывает r– и К–отбор, но r–отбор преобладает на ранней стадии развития популяции, а К–отбор характерен для стабилизированных систем, то есть r– стратегия обеспечивает выживание за счет количественного роста, а К–стратегия – за счет качественного совершенствования взаимоотношений между особями и особей с абиотической средой. Эти две стратегии по существу представляют два различных решения одной задачи – длительного выживания вида.
Таблица 9 – Характерные особенности r– и К– видов

Таким образом, виды с r–стратегией быстрее заселяют нарушенные местообитания (обнаженная горная порода, лесные вырубки), чем виды с К–стратегией, так как они легче распространяются и быстрее размножаются. Виды с К–стратегией более конкурентоспособны, и они обычно они вытесняют r–виды, которые тем временем перемещаются в другие нарушенные местообитания. Высокий репродуктивный потенциал r–видов свидетельствует о том, что оставшись в каком–либо местообитания, они быстро использовали бы доступные ресурсы и превысили поддерживающую емкость среды, а затем популяция погибла бы. Виды с r–стратегией занимают данное местообитание в течение жизни одного или, самое большее, нескольких поколений. В дальнейшем они переселяются на новое место. Отдельные популяции могут регулярно вымирать, но вид при этом перемещается и выживает. В целом эту стратегию можно охарактеризовать как стратегию «борьбы и бегства ».
Следует отметить, что одну и ту же среду обитания разные популяции могут использовать по–разному, поэтому в одном и том же местообитании могут сосуществовать виды с r– и К–стратегией. Между этими крайними стратегиями существуют переходы. Ни один из видов не подвержен только r– или только К–отбору. В целом же r– и К–стратегии объясняют связь между разнокачественными характеристиками популяции и условиями среды.
3.1.2. Видовая структура сообществ
В природе популяции разных видов объединяются в макросистемы более высокого ранга – так называемые сообщества или биоценозы. Термин «биоценоз» был предложен немецким гидробиологом Карлом Мебиусом в 1877 г.
Биоценоз (от греч. bios – жизнь, koinos – общий) – это совокупность популяций всех видов живых организмов, населяющих определенную географическую территорию, отличающуюся от других соседних территорий по химическому составу почв, вод, а также по ряду физических показателей (высота над уровнем моря, величина солнечного облучения и т.д.).
В природе биоценозы бывают разного масштаба. Это, например, биоценоз моховой кочки, разрушающегося пня, луга, пруда, болота, леса (рис. 105).
В состав биоценоза входят следующие компоненты (рис. 106):
фитоценоз – совокупность растений на определенной территории;
зооценоз – животные, проживающие в пределах фитоценоза;
микробоценоз – совокупность микроорганизмов, населяющих почву.
Конкретные сообщества складываются в строго определенных условиях окружающей среды – (почва, грунтовые воды, климат.). Взаимодействуя с компонентами биоценоза, почва и грунтовые воды образуют эдафотоп, а атмосфера – климатоп. Компоненты неживой природы, образуют косное единство – экотоп. Относительно однородное по абиотическим факторам среды пространство, занятое биоценозом называется биотопом. Это может быть какой–то участок суши или водоема, берег моря или склон горы. Биотоп это неорганическая среда, которая является необходимым условием существования биоценоза.

Рисунок 105 – Биоценоз дубравы

Рисунок 106 – Структура биоценоза и взаимодействия его компонентов
Биоценоз имеет видовую, пространственную и экологическую структуры.
Видовая структура биоценоза – это разнообразие в нем видов и соотношение их численности или массы. Различают понятия «видовое богатство» и «видовое разнообразие» биоценозов. Видовое богатство – это общий набор видов сообщества, который выражается списками представителей разных групп организмов. Видовое разнообразие – это показатель, отражающий не только качественный состав биоценоза, но и количественные взаимоотношения видов.
Видовое богатство и видовая насыщенность зависят от многих факторов, в первую очередь от условий существования фитоценоза. Главными лимитирующими факторами видового разнообразия являются температура, влажность и наличие пищевых ресурсов. Чем богаче, то есть чем более благоприятны по сочетанию вышеуказанных факторов, условия местообитания, тем разнообразней видовой состав и продуктивность компонентов биогеоценоза. Чем сильнее для большинства видов условия произрастания отклоняются от оптимума, тем беднее и менее продуктивно сообщество, но зато тем характернее для него слагающие его биологические виды и тем многочисленней семьи организмов этих видов (Гиляров, 1980).
Самые богатые по видовому составу биоценозы тропических лесов, с разнообразным животным миром. В них трудно найти даже два рядом стоящих дерева одного вида. Наиболее бедны видами биоценозы (сообщества) экосистем высоких широт, пустынь и высокогорий. Здесь могут выжить организмы, жизненные формы которых приспособлены к таким условиям. Например, в биоценозы тропических лесов входят десятки тысяч видов растений, сотни тысяч видов беспозвоночных и несколько тысяч видов позвоночных животных. Биоценозы же тундры, пустыни включают несравненно меньшее количество видов. Так, на Таймыре в тундровом биоценозе насчитывается всего 139 видов высших растений, 670 видов низших растений, около 1000 видов животных и 2500 видов микроорганизмов. Естественно, их биомасса и продуктивность в десятки и сотни раз меньше, чем в лесах тропиков и субтропиков.
Условия произрастания могут сильно различаться по теплообеспеченности, влажности и богатству почв в пределах одного региона – в зависимости от положения в рельефе и почвообразующей породы. Поэтому, видовое разнообразие фитоценозов разных экотопов может различаться в десятки раз. Например, на севере Дальнего Востока оптимальные для региона экотопы формируются в поймах реки на южных склонах гор, на юге – в средней части северных теневых склонов. Наименее благоприятные экотопы свойственны соответственно переувлажненным с наличием вечной мерзлоты в почвенном профиле северным склонам, шлейфам склонов и низменностям, на юге – крутым сухим южным склонам с маломощными каменистыми почвами и заболоченным низменностям с водоупорными глинами под почвенным слоем. В результате в поймах рек Магаданской области растут самые сложные по видовому составу высокопродуктивные травяные лиственничники (до 40–50 видов), а на северных склонах гор Южного Приморья – уникальные дальневосточные хвойно–широколиственные леса (70–80 видов). В наиболее суровых условиях на севере Дальнего Востока формируются самые бедные по составу (не более 20 видов) низко продуктивные монодоминантные леса – заболоченные лиственничники осоково–сфагновые, на юге – сухие дубняки осоковые и марьянниково–осоковые.
Видовое разнообразие взаимосвязано и с разнообразием условий конкретной среды обитания, т.е. в пределах экотопа. Чем больше организмов найдут в данном биотопе подходящих для себя условий по экологическим требованиям, тем больше видов в нем поселится.
Есть и другое обстоятельство, определяющее видовую насыщенность и видовое богатство: возраст данного фитоценоза, т. е. продолжительность времени, в течение которого растения, входящие в состав фитоценоза, сживались друг с другом. Молодые, только начинающие развиваться ценозы – бедны видами по сравнению со зрелыми, или климаксными, сообществами. Чем «старше» фитоценоз, тем при прочих равных условиях большее число видов входит в его состав, а чем «моложе» – тем меньше. А возраст фитоценоза в свою очередь зависит от возраста той территории (или акватории), где он расположен.
Сказывается и воздействие человека, и степень изменения среды самим сообществом (максимальная в лесах и минимальная в пустынях) В агроценоз пшеничного поля, кроме пшеницы, входят, хотя бы в минимальном количестве, разнообразные сорняки, насекомые–вредители пшеницы и хищники, питающиеся фитофагами, мышевидные грызуны, беспозвоночные – обитатели почвы и напочвенного слоя, микроскопические организмы ризосферы, патогенные грибки и многие другие.
Каждый конкретный биоценоз характеризуется строго определенным видовым составом. При этом одни виды биоценоза могут быть представлены многочисленными популяциями, а другие малочисленными. В связи с этим в любом биоценозе можно выделить один или несколько видов, определяющих его облик. Эти виды называются доминирующими или доминантами. Они занимают ведущее, господствующее, положение в биоценозе. В лесу, который состоит из десятков видов растений, обычно один или два дают до 90 % древесины. Обычно наземные биоценозы называют по доминирующим видам: лиственничный лес, дубрава или дубняк, сфагновое болото, ковыльно–типчаковая степь. Чем бедней видовой состав, тем больше видов доминантов.
Виды зооценоза, живущие за счет доминантов, получили название предоминантов. К примеру, в дубовом лесу таковыми являются кормящиеся на дубе насекомые, сойки, мышевидные грызуны.
Между численностью видов–доминантов и общим видовым богатством сообщества существует определенная связь. Со снижением числа видов обычно резко повышается обилие отдельных форм. В таких обедненных сообществах ослабевают биоценотические связи и некоторые наиболее конкурентоспособные виды получают возможность беспрепятственно размножаться.
Чем специфичнее условия среды, тем беднее видовой состав сообщества и тем выше может быть численность отдельных видов. Эта закономерность получила название правила А. Тинемана, по имени немецкого ученого, изучавшего особенности видовой структуры сообществ в 30–е годы прошлого века. В бедных видами биоценозах численность отдельных видов может быть чрезвычайно высокой. Достаточно вспомнить вспышки массового размножения леммингов в тундрах или насекомых–вредителей в агроценозах (рис. 107). Подобную закономерность можно проследить в сообществах самого разного масштаба. В буртах свежего конского навоза, например, почти анаэробная обстановка, много аммиака и других токсичных газов, высокая температура за счет деятельности микроорганизмов, т. е. создаются резко специфичные, отклоняющиеся от обычной нормы условия жизни для различных животных. В таких буртах видовой состав беспозвоночных вначале крайне беден. Развиваются личинки мух–дрозофил, и размножаются немногие виды нематод–сапрофагов (семейство Rhabditidae) и хищных гамазовых клещей (род Parasitus). Но зато все эти виды чрезвычайно многочисленны, редких форм почти нет. В подобных случаях кривая, описывающая распределение видов по их численности, имеет сильно сглаженную левую часть. Такие сообщества неустойчивы и отличаются резкими колебаниями обилия отдельных видов.
Рисунок 107 – Структура доминирования в сообществе насекомых стеблестоя зерновых на полях (по Н. И. Куликову, 1988). На оси абсцисс – виды в порядке убывания численности
Постепенно, по мере разложения навоза и смягчения условий среды, видовое разнообразие беспозвоночных нарастает, при этом заметно снижается относительная и абсолютная численность массовых форм.
В наиболее богатых биоценозах практически все виды малочисленны. В тропических лесах редко можно встретить рядом несколько деревьев одной породы. В таких сообществах не происходит вспышек массового размножения отдельных видов, биоценозы отличаются высокой стабильностью. Кривая, отражающая видовую структуру такого типа, имела бы на рисунке 108 особенно крутую левую часть.
Таким образом, даже самый общий анализ видовой структуры может дать достаточно много для целостной характеристики сообщества. Разнообразие биоценоза тесно связано с его устойчивостью. Деятельность человека сильно сокращает разнообразие в природных сообществах. Это вызывает необходимость предвидеть ее последствия и принимать меры для поддержания природных систем.
Среди доминантов есть такие, без которых другие виды существовать не могут. Их называют эдификаторами. Эдификаторам (от лат. – строители) принадлежат главные средообразующие функции фитоценоза. Они определяют микросреду (микроклимат) всего сообщества и их удаление грозит полным разрушением биоценоза. Это виды, создающие условия для жизни других видов, ими во многом определяются особенности биоценоза. Эдификаторы всегда доминанты, но доминанты не всегда эдификаторы. Как правило, эдификаторами служат растения – ель, сосна, ковыль, и лишь изредка – животные (сурки). На территориях занятых колониями сурков, именно их роющая деятельность определяет большей частью характер ландшафта, микроклимат и условия произрастания растений.
Сообщества разного состава эдификаторов различаются продуктивностью, степенью проявления средообразующих функций, хозяйственной значимостью, устойчивостью по отношению к воздействиям внешних факторов. Эдификаторов обычно немного. Например, в зависимости от породного состава эдификаторного древесного яруса, лесные биогеоценозы могут быть сосновыми, березовыми, березово–еловыми (еловыми с примесью березы), елово–сосново–березовыми (березовыми с примесью ели и сосны) или елово–березовыми. Эдификаторную роль в таких биогеоценозах выполняют соответственно сосна, береза, ель вместе с березой, береза с участием сосны и ели, береза с участием ели (рис. 108).

Рисунок 108 – Структура доминирования в сообществе насекомых стеблестоя зерновых на полях (по Н. И. Куликову, 1988). На оси абсцисс – виды в порядке убывания численности
В сообществах нередко несколько пород, выполняющих большие средообразующие функции, одна или две из них являются главными эдификаторами, а остальные несколько уступают главным. Они – соэдификаторы.
Господствующие виды во второстепенных ярусах и обладающих сильными эдификаторными свойствами являются субэдификаторами. К ним относятся кедровый стланик, образующий густой подлесок в северных лиственничниках, брусника и зеленые мхи в бруснично–зеленомошных типах леса, сфагнум в редколесьях сосны и лиственницы на Крайнем Севере, вейник Лангсдорфа в травяных типах леса и др.
Виды–эдификаторы обязательны для любого биоценоза. Однако роль их в тех или биоценозах не абсолютна. В таежных экосистемах при изреживании леса ель может утратить функции эдификатора, поскольку при этом происходит осветление леса и в него внедряются другие породы. А в заболоченных еловых редколесьях со сплошным моховым покровом главным эдификатором выступают сфагновые мхи. Сфагнум является очень сильным средообразователем. Он определяет специфические условия биоценоза. Почвы в этих редколесьях отличаются плохой аэрацией и низкой теплопроводностью торфа, кислой реакцией среды, бедностью элементов минерального питания для высших растений. В степных биоценозах мощным эдификатором, подстать сфагнуму, служит ковыль.
«Второстепенные» виды – малочисленные и даже редкие тоже очень важны в сообществе. Их присутствие – это гарантия устойчивого развития сообществ.
Ассектаторы – оказывают малое влияние на среду внутри сообщества. Это виды–спутники, большинство из них – второстепенные по представленности в ценозе. Как правило, у каждого вида, обладающего сильными средообразующими функциями, т.е. у мощных эдификаторов, формируются собственные свиты видов–спутников. Таковы многие виды в каменноберезняках, в кедрово–широколиственных лесах, в кедровостланиковых зарослях. Например, виды–спутники кедра корейского: орех маньчжурский, бархат амурский, диморфант, ясень носолистный, ильм лопастной, клены зеленокорый, ложнозибольдова, маньчжурский, граб сердцелистный, многие кустарники: лещина, чубушник, представители сем. Аралиевых, из трав: разнообразные папоротники и крупнотравье, тригонотис укореняющийся, кислица обыкновенная, аризема мощная и амурская, лесной мак весенний, разные виды хохлаток и ветрениц и многие другие виды.
Показатели численности видов и их динамика являются одними из основных в изучении биоценоза. Численность определяется визуально и инструментально, но чаще визуально. Всегда на учетной единице: площади (дм, м2, км2, га), длины (м, км), объема (м3, 10 дм3), времени (час, сутки) и т.д.
Как известно, видовая структура биоценоза характеризуется не только числом видов, входящих в его состав, т.е. видовым разнообразием, но и их количественными показателями. Например, если в двух сравниваемых биоценозах растения представлены двумя видами и в каком–то из них 90 % особей принадлежит одному виду, тогда как в другом на долю особей каждого вида приходится по 50 %, то эти биоценозы будут сильно отличаться друг от друга.
Количественное соотношение видов в биоценозе, или индекс разнообразия (Н) чаще всего определяется по формуле Шеннона:
H = – ?pi*log2pi,
где pi – доля каждого вида в сообществе
Встречаемость (частота встречаемости, коэффициент встречаемости) – это относительное число выборок, в которых встречается вид. Если выборка состоит из 100 учетных площадок, а вид отмечен на 43, то и встречаемость равна 43 %. При встречаемости 25%, вид встречается в каждой четвертой площадке учета и он случайный. Высокая встречаемость, если вид отмечен более чем на 50 % учетных площадках. Обычно закладывается 50 учетных площадок, но не менее 25.
Покрытие – процент площади, покрываемой надземными частями растений. Процент площади, занятой основаниями растений – истинное покрытие, верхними частями – проективное. Проективное покрытие – обязательный показатель при изучении напочвенного покрова.
При изучении древесно–кустарниковых ярусов синонимом проективного покрытия служит сомкнутость – отношение площади проекций крон к площади занимаемого участка; в отличие от проективного покрытия сомкнутость измеряется в долях от единицы. Истинное проективное покрытие для древостоя – сумма площадей поперечного сечения стволов и полнота, определяется расчетным путем по данным перечета древостоя.
Биомасса – общие запасы органического вещества, накопленные к моменту учета. Выражаются в массе абсолютно–сухого, воздушно–сухого или сырого вещества. Биомасса растений – растительная масса, фитомасса; биомасса животных – зоомасса. Биомасса, ее фракционная структура, скорость накопления (продукция – прирост биомассы за определенный промежуток времени) являются важнейшими – интегральными, показателями жизнедеятельности организмов. Они дают возможность оценить роль каждого фактора и популяции в формировании биогеоценоза, оценить запасы биологических и пищевых ресурсов, сделать кратко– и долгосрочные прогнозы развития сообществ, предсказать пути их трансформации и разработать мероприятия по охране и рациональному использованию любого из ресурсов.
Для оценки видового разнообразия используют и другие показатели.
Обилие – это количество особей вида на единице площади или объема. Наиболее часто используются шкалы обилия Друде и Хульта
Постоянство – отношение числа содержащих изучаемый вид выборок к общему числу выборок, выраженное в процентах. В зависимости от величины этого отношения различают следующие категории видов: постоянные – встречаются более чем в 50% выборок, добавочные – в 25–50% выборок, случайные – менее чем в 25% выборок.
Степень доминирования – отношение (обычно в процентах) числа особей данного вида к общему числу всех особей рассматриваемой группировки. Так, если из 200 птиц, зарегистрированных на данной территории, 100 составляют зяблики, степень доминирования этого вида среди птиц составляет 50 %. На территориях занятых колониями сурков, именно их роющая деятельность определяет большей частью характер ландшафта, микроклимат и условия произрастания растений.
Наиболее благоприятные условия для существования множества видов характерны для переходных зон между сообществами, которые называют экотонами, а тенденцию к увеличению здесь видового разнообразия называют краевым эффектом. Экотон богат видами, прежде всего потому, что они попадают сюда из пограничных сообществ, но, кроме того, он может содержать и свои характерные виды, которых нет в этих сообществах. Яркий пример: лесная опушка, на которой пышнее и богаче растительность, гнездится значительно больше птиц, больше насекомых, чем в глубине леса. Здесь разнообразнее условия освещенности, влажности, температуры. Чем сильнее различия двух соседствующих биоценозов, тем разнороднее условия на их границах и тем сильнее проявляется пограничный эффект.
В пределах любого биогеоценоза всегда можно обнаружить различные группировки организмов, приуроченных к какому–либо одному виду, без которого они не смогут существовать. Чаще всего это объединение происходит на основе популяции, занимающей важное средообразующее (эдификаторное) положение в сообществе. Например, ель в еловом лесу, занимая доминирующее положение, обусловливает сосуществование целого комплекса растений и животных, которые в своей жизнедеятельности тесно с ней связаны. Это кислица, многие грибы, белки, птицы (пищуха, поползень, клест–еловик), короеды и многие другие. Все они в своих проявлениях адаптировались к существованию во взаимосвязи с елью. На ней они находят питание, там же строят гнезда, укрываются от врагов и неблагоприятной погоды. Такие группировки разнородных организмов, тесно объединенных в своей жизнедеятельности вокруг одного какого–то вида в сообществе, называют консорцией (лат. consortio — общение, соучастие). Ель, таким образом, в биогеоценозе является отдельной консорцией вместе со всеми ее обитателями и посетителями, так же как береза со всей своей свитой консортов или осина, дуб, сосна.
Впервые термин «консорция» был введен Л. Г. Раменским в 1952 году для обозначения группы видов, взаимосвязанных между собой в биогеоценозе. Почти одновременно с Л. Г. Раменским, в том же 1952 году, сходное представление о консорциях сформулировал В. Н. Беклемишев. Л. Г. Раменский писал, что помимо цепей питания «целесообразно выделить в ценозах также сочетание разнородных организмов, тесно связанных друг с другом в их жизнедеятельности известной общностью их судьбы (консортивные группы, или консорции). Такова, например, древесная порода (ель, береза, липа, дуб) со свойственными ей паразитами, сапрофитами, эпифитами (лишайники, мхи), симбионтами (микориза, микробы ризосферы и др.), вредителями – переносчиками пыльцы, семян и т. д.». Обычно консорция формируется на базе популяции автотрофного растения (ель, осина, береза, ковыль и др.). В этом случае его называют детерминантом, а виды, объединенные вокруг него, – консортами.
Среди консортов имеются виды, получающие от детерминанта питание и энергию, то есть связанные с ним трофически (пищевыми связями), и есть виды, связанные топически (находящие на нем укрытие и жилье), то есть располагающиеся на нем. Например, различные фитотрофы (листоеды, поедающие древесину, плоды или пьющие соки) связаны с детерминантом трофически, тогда как эпифиты, хищники (птицы, насекомые) связаны топически. Нередки также виды, связанные с детерминантом трофически и топически, то есть питающиеся его веществами и на нем же строящие свое жилье. Группы консортов того или иного порядка, объединенные вокруг детерминанта, по предложению В. В. Мазинга названы концентрами.
Многие консорты одновременно и сами являются детерминантами других консорций. Например, белка, будучи консортом ели, имеет своих консортов из хищников (куница), паразитов, располагающихся на ее теле или в ее гнезде, и организмов, питающихся ее выделениями и пометом. Входя в консорцию ели, белка вносит в нее свою свиту (своих консортов). В связи с этим все виды в консорции можно разделить на консортов первого порядка, консортов второго порядка. В таблице 4 представлена взаимозависимость детерминанта и его консортов, а также распределение консортов по концентрам I – III порядков.
Таблица 10 – Консорция дуба обыкновенного (по Т.А. Работнову, 1974)
Как видим, каждая консорция охватывает большое количество видов. Число их велико еще и потому, что детерминанты представлены в биогеоценозах разновозрастным составом, причем нередко каждой возрастной стадии организмов сопутствует свое консортивное население. Например, стволовые вредители поражают главным образом взрослые ели, а ржавчинные грибы – молодые экземпляры. С возрастом ели меняется состав ее консортов, питающихся корневыми выделениями, а также микоризообразующих грибов.
Среди консортов имеются виды, которые входят в состав только одной консорции, а также двух и более консорций, способствуя тем самым объединению организмов биогеоценоза в единый комплекс. Связи в консорции могут быть длительные, на протяжении всей жизни консортов (тут они размножаются и выводят потомство – жуки–короеды, усачи; растут, развиваются – грибы–паразиты); или носить сезонный характер (хвоееды, клест–еловик и др.).
Состав видов в консорции – результат длительного коэволюционного процесса, происходящего по мере становления автотрофного растения и подбора видов, консортивно связанных с ним и способных существовать в условиях местообитания детерминанта.
Каждая консорция представляет собой особую структурную единицу биогеоценоза. В биогеоценозе консорций может быть мало, даже одна (например, чистый ельник, березняк) и много (например, смешанный лес, злаково–полынные степи, влажные тропические леса и др.). Выпадение детерминанта из биогеоценоза сопровождается выпадением и многих его консортов, особенно монофагов, то есть питающихся исключительно за счет организмов данной консорции. Виды, входящие в несколько консорций, обычно остаются в биогеоценозе. Наоборот, внедрение детерминанта в биогеоценоз может сопровождаться включением и его консортов, особенно если детерминант обычен для данной местности. В том случае, когда он интродуцирован из отдаленных районов (например, специально или случайно ввезен человеком), консорция создается из местных видов. Нередко интродуцированный в биогеоценоз вид существует без консортивных связей. Не поедаемый никем, он при этом чрезвычайно сильно увеличивает численность, влияет на структуру биогеоценоза и вызывает в нем катастрофические изменения.
Изучение закономерностей состава и взаимосвязей между консортами, является в настоящее время предметом пристального внимания биогеоценологов. Их исследования важны не только для теории, но и для практики регулирования природных процессов (борьба с вредителями, интродукция новых видов и охрана природных комплексов, охотоведение и рыбное хозяйство и т.д.), и для моделирования биогеоценозов. Изучение консортивных связей в биогеоценозе позволяет полнее охарактеризовать его структуру, насыщенность населения и многообразие биотических связей.
|