Тема 3.2. Трофическая структура сообществ
Особи разных видов существуют в биоценозах не изолированно; они вступают между собой в разнообразные прямые и косвенные отношения. Экологическое взаимодействие популяций имеет цель предельную синхронизацию жизнедеятельности всех популяций в биоценозе и обеспечение круговорота биогенных элементов в экосистеме. Согласно классификации В.Н. Беклемищева (1951), межвидовые отношения по значению, которое они имеют для занятия видом в биоценозе определенного положения подразделяются на 4 типа: - трофические,
- топические,
- форические,
- фабрические.
Наиболее важными являются трофические и топические связи, так как именно они удерживают организмы разных видов друг возле друга, объединяя их в сообщества. Трофические связи возникают между видами, когда один вид питается другим: живыми особями, мертвыми остатками, продуктами жизнедеятельности. Трофическая связь может быть прямой и косвенной. Прямая связь проявляется при питании львов живыми антилопами, гиен трупами зебр, жуков–навозников пометом крупных копытных, пчел, нектаром растений, божьей коровки, тлей, коровы травой и т.д. Насекомоядные птицы не питаются растениями, но поедают многих насекомых, которые кормятся листьями или опыляют цветки. Поедая насекомых–опылителей, птицы косвенным образом воздействуют на число производимых деревом плодов, то есть на количество пищи, доступной животным, которые питаются плодами и проростками, на хищников и паразитов этих животных и т.д. Топические связи характеризуют любое физическое или химическое изменение условий обитания одного вида в результате жизнедеятельности другого. Эти связи крайне разнообразны. Они заключаются в создании одним видом среды для другого. Морские желуди, поселяющиеся на коже китов, личинки мух, обитающие в лепешках коровьевого навоза, лишайники на стволах деревьев связаны прямой топической связью с теми организмами, которые предоставляют им субстрат или среду обитания. Ель, затеняя почву, вытесняет светолюбивые виды из–под своей кроны. Почвенные микроорганизмы формируют почву – среду обитания многих видов растений и животных, улучшая ее структуру и обогащая химический состав. Водные зеленые растения (водоросли, фитопланктон) выделяют в воду кислород, создавая условия существования консументам. Лесные древесные растения создают возможность существования многим видам насекомых, птиц, животных, не только обеспечивая их кормом, но и создавая особый микроклимат, места для поселения, охоты, защиты и т.п. Все популяции экосистемы в первую очередь доминирующие формируют экоклимат – совокупность температурных, световых, влажностных и иных параметров экотопа, и, одновременно, зависят от него, т.е. находятся друг с другом в прямых или косвенных топических связях. Форические связи – это участие одного вида в распространении другого. В роли транспортировщиков выступают животные. Перенос животными семян, спор, пыльцы растений называют зоохорией, перенос других, более мелких животных – форезией (от лат. форас – наружу, вон) (жуки–навозники – клещи: нередко ползают с поднятыми подкрыльями, так как не в состоянии их сложить из–за густо усеявших тело клещей.). Перенос осуществляется обычно с помощью специальных и разнообразных приспособлений. Животные могут захватывать семена растений двумя способами: пассивным и активным. Пассивный захват происходит при случайном соприкосновении тела животного с растением, семена или соплодия которого обладают специальными зацепками, крючками, выростами (череда, лопух). Распространителями их обычно служат млекопитающие, которые на шерсти переносят такие плоды иногда на довольно значительные расстояния. Активный способ захвата – поедание плодов и ягод. Не поддающиеся перевариванию семена животные выделяют вместе с пометом. Фабрические связи – это такой тип биоценотических отношений, в который вступает вид, используя для своих сооружений (фабрикаций) продукты выделения или мертвые остатки или даже живых особей другого вида. Например, птицы для постройки гнезд используют ветви деревьев, листья, траву, шерсть млекопитающих, пух и перья, личинки ручейников для строительства своих домиков используют кусочки коры, песчинки, обломки раковин или же сами раковины с живыми моллюсками мелких видов. Существование сообщества предполагает благоприятное сочетание большинства экологических факторов. В связи с тем, что оптимум почти недостижим, принято различать: физиологический оптимум – оптимальное для вида сочетание всех видов абиотических факторов, при котором возможны наиболее быстрые темпы роста и размножения; синэкологический оптимум – оптимальное сочетание биотических факторов, при котором вид испытывает наименьшее давление со стороны врагов и конкурентов, что позволяет успешно ему размножаться. Физиологический и синэкологический оптимумы не всегда совпадают. Причем вид в подавляющем большинстве случаев живет в синэкологическом, а не в физиологическом оптимуме. Например, массовое размножение гессенской мухи – вредителя зерновых – после особенно суровых зим, которые должны были бы неблагоприятно сказаться на численности данного насекомого. В нормальные по условиям годы гессенскую муху сильно истребляют несколько видов ее естественных врагов – паразитических перепончатокрылых наездников. Из–за слабой морозоустойчивости в суровые зимы враги гессенской мухи погибают практически полностью. Это дает возможность вредителю восстановить собственную численность, уменьшенную низкими температурами в зимний период, и беспрепятственно размножаться в угрожающем для урожая количестве. Многие галофиты лучше развиваются не на засоленных почвах, где они образуют сообщества, а на влажных почвах с незначительным содержанием солей. Многие растения скал с ксероморфным обликом свой экологический оптимум имеют на лугах. Несовпадение экологического и физиологического оптимумов – результат борьбы за существование между растениями. В ряде случаев в процессе борьбы за существование растения оттесняются в экстремальные условия из более благоприятных для них фитоценозов. Например, пихта белокорая и ель саянская не потому растут в более высоких горных поясах, что там лучше условия, а потому, что туда их вытесняют ель корейская, кедр и пихта цельнолиствная. Так же и светлолюбивые осина и береза уступают свои более благоприятные экотопы темнохвойным породам. Точно также травами из пойменных местообитаний вытесняются мхи и кустарнички. Межвидовые связи, формирующие биоценоз, обусловливают закономерные соотношения в нем видов, их экологических особенностей, численности, распределения в пространстве, т.е. создают определенную структуру биоценоза. Разнообразие возможных биотических межвидовых взаимоотношений представлено на рисунке 109. 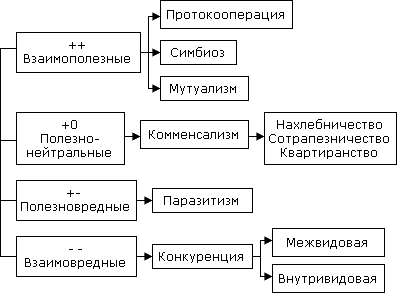
Рисунок 109 – Основные типы биотических межвидовых взаимоотношений
Таким образом, все биотические связи можно разделить на 6 групп: популяции не влияют друг на друга (00); популяции имеют взаимовыгодные связи (++); отношения вредны для обоих видов (– –); один из видов получает выгоду, другой испытывает угнетение (+–); один вид получает пользу, другой не испытывает вреда (+0); один вид угнетается, другой не извлекает пользы (– 0); Причем, несмотря на многообразие, их можно разделить на две основные группы – положительные и отрицательные. Положительные взаимодействия в биоценозе. Симбиоз – система отношений, при которой формируются тесные функциональные взаимодействия, выгодные для обоих видов (мутуализм) или только для одного из них (комменсализм). Комменсализм – взаимоотношения, при которых один из партнеров получает пользу, не нанося ущерба другому (усоногие рачки, прикрепляющиеся к коже кита, преимущество – более быстрое передвижение). Различают три формы комменсализма: - основанный на потреблении остатков пищи хозяев, называют трофобиоз или нахлебничество. Таковы, например, взаимоотношения львов и гиен, подбирающих остатки недоеденной пищи, или акул с рыбами–прилипалами.
- синойкия (квартиранство). Примером синойкии являются взаимоотношения некоторых актиний и тропических рыбок, которые укрываются от нападения хищников среди щупалец актиний.
- сотрапезничество – потребление разных веществ или частей одного и того же ресурса.
Например, взаимоотношения между различными видами почвенных бактерий, перерабатывающих разные органические вещества из перегнивших растительных остатков и высшими растениями, которые потребляют образовавшиеся при этом минеральные соли. Комменсализм обычно связан с поиском пищи или необходимого укрытия. Примером комменсализма является сосуществование некоторых мелких рыб с крупными актиниями (рис. 110). Рыбки нечувствительны к стрекательным нитям актинии и «подбирают» остатки пищи между щупальцами, а актинии невольно обеспечивают этим рыбкам защиту. Очень часто комменсалами являются бактерии и простейшие, живущие в кишечнике и на поверхности более крупных организмов.

Рисунок 110 – Примеры комменсализма. Слева направо: рыба–клоун и актиния, цапли и буйвол
Наиболее характерные растения, которые можно привести в качестве примеров комменсализма по способу размещения в ценозе и по типу питания: эпифиты, лианы, почвенные и наземные сапрофиты. Эпифиты – растения, как высшие, так и низшие, растущие на других (хозяевах): деревьях, кустарниках, которые служат ему опорой. Отношения эпифитов к их хозяевам могут быть определены как комменсализм, при котором один из вступающих в эти отношения видов получает какое–либо преимущество, а второй не терпит ущерба. Преимущество при этом получает эпифит. Чрезмерное развитие эпифитов на стволах и ветвях может угнетать и даже быть причиной поломки ствола растения–хозяина. Эпифиты могут затруднять рост и ассимиляцию, а также способствовать загниванию тканей хозяина вследствие повышения влажности. В зависимости от условий существования эпифиты (Ричардс, 1961) делятся на три группы: теневые, солнечные и крайне ксерофильные. Теневые эпифиты обитают в условиях сильной затененности, малого и мало меняющегося дефицита насыщения, то есть в условиях, почти не отличающихся от условий жизни наземных трав. Они обитают главным образом в третьем (нижнем) ярусе леса. Многие из них имеют гигроморфную структуру тканей. Группа солнечных эпифитов, наиболее богатая по числу видов и особей, связана с кронами деревьев верхних ярусов. Эти эпифиты живут в микроклимате, промежуточном между микроклиматами напочвенного покрова и открытых мест, и получают гораздо больше света, чем теневые эпифиты. Многие солнечные эпифиты более или менее ксероморфны; осмотическое давление у них выше, чем у теневых эпифитов. Крайне ксерофильные эпифиты живут на верхушечных ветвях более высоких деревьев. Условия их местообитания подобны условиям открытых мест, условия питания здесь крайне суровы. Эпифиты, как правило, сапротрофы, то есть, питаются за счет отмирающих тканей растения–хозяина (рис. 124). Обычно для разложения этих отмирающих тканей эпифиты используют грибы, образующие микоризу с корнями эпифита. Большую роль в питании играют некоторые животные. Так, муравьи, поселяясь среди корней эпифитов, приносят в свои гнезда большое количество мертвых листьев, семян, плодов, которые, разлагаясь, дают питательные вещества эпифитам. Некоторые беспозвоночные и позвоночные животные поселяются в воде, скапливающейся в чашах, образованных листьями эпифитов из семейства бромелиевых, и их трупы, разлагаясь, дают пищу эпифитам. Наконец, среди эпифитов есть и насекомоядные растения, например виды рода непентес (Nepenthes) и некоторые пузырчатки.

Рисунок 111 – Эпифиты–сапротрофы – зеленые мхи на дереворазрушающем грибе в лиственничнике Магаданской области
Влажные тропические леса богаты эпифиллами–эпифитами, обитающими на листьях растений. Их существование связано с долговечностью вечнозеленых листьев, а также с высокой влажностью и температурой среды. Эпифиллы обитают чаще всего на листьях невысоких деревьев, иногда на листьях травянистых растений. К эпифиллам относятся водоросли, лишайники, печеночники; эпифильные листостебельные мхи редки. Иногда наблюдаются эпифиллы, растущие на эпифиллах, например водоросль, растущая на эпифильном мхе. Лианы. К лианам относят высшие растения со слабыми стеблями, нуждающимися в какой–нибудь опоре, чтобы подняться вверх. Лианы, относятся к комменсалам, но изредка могут причинить ущерб и даже вызвать гибель деревьев. Лианы делят на две группы: мелкие и крупные. Среди мелких лиан преобладают травянистые формы, хотя есть и деревянистые. Они развиваются в нижних ярусах лесов, а иногда (вьюнок – Convolvulus, подмаренник – Galium, марена – Rubia, княжик – Clematis, и др.) и среди травяного покрова. Крупные лианы обычно бывают деревянистыми. Они достигают вершин деревьев второго, иногда первого яруса. У этих лиан обычно очень длинные и иногда настолько крупные водоносные сосуды, что они видны на поперечном срезе простым глазом. Эта особенность связана с необходимостью поднимать огромные количества воды в крону лианы, иногда не уступающую по величине кроне дерева, по стволу, диаметр которого во много раз меньше диаметра обычного дерева. Стебли лиан часто имеют очень длинные междоузлия и быстро растут не ветвясь, пока не достигнут того яруса, в котором обычно развертывается листва этих растений. В «уссурийской тайге» длина взрослых лиан актинидии и винограда амурского достигает нескольких десятков метров, а диаметр – 10 и более сантиметров. Крупные лианы иногда так быстро растут и развиваются в таких массах, что губят поддерживающие их деревья. Вместе с деревом–опорой лиана падает на землю и здесь гибнет или поднимается на другое дерево. Часто расстояние между основаниями стволов лианы и дерева–опоры измеряется десятком или несколькими десятками метров, что убеждает в том, что несколько промежуточных деревьев, служивших лиане опорой, погибли ранее. Часто лианы фестонами перекинуты с одного дерева на другое, достигая длины 70, а в исключительных случаях (ротанговые пальмы) 240 м. В лесах умеренного пояса распространены исключительно или почти исключительно мелкие лианы, поэтому они не играют здесь большой роли. Почвенные и наземные сапрофиты. Сапрофиты – растительные организмы, живущие полностью (полные сапрофиты) или частично (частичные сапрофиты) за счет отмерших органов животных и растений. Кроме эпифитов, которые относятся по способу питания к сапрофитам, в эту группу входят многие наземные растения и обитатели почвы. К сапрофитам относится большинство грибов и бактерий, играющих огромную роль в круговороте веществ в почве, а также некоторые цветковые растения из семейств орхидных (гнездоцветка клобучковая) и вертляницевых (вертляница одноцветковая) в лесах умеренного пояса и из семейств лилейных, орхидных, горечавковых, истодовых и некоторых других в лесах тропического пояса. Большинство этих цветковых растений – полные сапрофиты, некоторые, по крайней мере, орхидеи, содержат какое–то количество хлорофилла и, вероятно, частично способны к фотосинтезу. Цвет надземных частей этих растений белый, светло–желтый, розовый, голубой или пурпурный. Обитают сапрофиты из цветковых растений в тропиках в тенистых местах на почве или на лежащих мертвых стволах. Обычно эти растения связаны с микоризными грибами, живущими на их корнях. Как правило, они невысоки, обычно не превышают 20 см, за исключением сапрофитной тропической орхидеи галеоны высочайшей (Gualala altissimo), представляющей собой лазающую (при помощи корней) лиану, достигающую высоты 40 м. Мутуализм – симбиотические взаимоотношения, неразделимые взаимополезные связи двух видов, предполагающие обязательное тесное сожительство организмов. Например, лишайник, сожительство гриба и водоросли – возник из паразитизма гриба на водорослях (рис. 112).
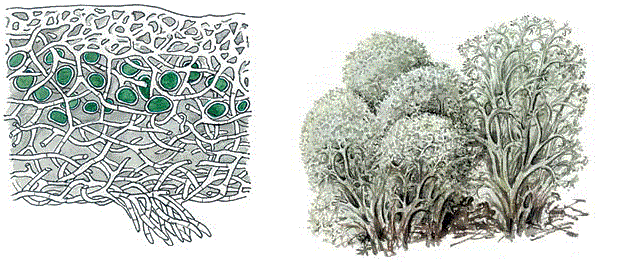
Рисунок 112 – Лишайник кладония – пример симбиоза. На рисунке: клетки водорослей – зелёного цвета; бесцветные – грибные гифы
Среди известных примеров симбиоза можно привести лишайники, сожительство рака–отшельника и актинии, симбиоз бактерий, переваривающих целлюлозу, и жвачных, взаимоотношения муравьев и тлей, которых они «пасут», получая взамен сладкие продукты выделения, рыбы–«санитары» и птицы–«санитары», уничтожающие паразитов на коже крупных млекопитающих и рыб (рис. 113). Важными примерами симбиоза является совместное существование грибов и деревьев, насекомых и цветковых растений.

Рисунок 113 – Симбиоз: Слева направо: мурена и креветка, муравьи и тли
К мутуализму относятся симбиоз азотфиксирующих организмов с голосеменными и цветковыми растениями – отношения между высшим растением и бактериями. На корнях многих растений встречаются клубеньки, образованные бактериями или реже грибами. Клубеньковые бактерии фиксируют атмосферный азот и переводят его в доступную для высших растений форму. Например, клубеньки на корнях растений из семейства бобовых образованы бактериями из рода ризобиум (Rhyzobium), а также на корнях видов лисохвоста, лоха, облепихи, подокарпуса, ольхи (Actinomyces alni) и других растений. Благодаря этому растения, зараженные клубеньковыми бактериями, могут хорошо расти на почвах, бедных азотом, а содержание азота в почве после культуры таких растений возрастает. В свою очередь бактерии получают от высших растений углеводы. Одной из разновидностей мутуализма является микориза – симбиотические отношения между высшим растением и грибом. Микоризы широко распространены среди диких и культурных растений. В настоящее время микориза известна более чем для 2000 видов высших растений (Федоров, 1954), но, несомненно, действительное число видов, для которых характерна микориза, значительно больше. Для высших растений, на корнях которых поселяются грибы, характерен особый тип питания – микотрофный. При микотрофном питании с помощью симбиотических грибов высшее растение получает зольные элементы пищи, в том числе азот, из органического вещества почвы. Что касается грибов, образующих микоризу, то они в большинстве не могут существовать без корневых систем высших растений, всасывающих влагу из почвы и поставляющих из кроны органическое вещество. Деревья растут гораздо лучше с микоризой, чем без нее. Различают два основных вида микоризы: эктотрофную и эндотрофную. При эктотрофной микоризе корень высшего растения окутывается плотным грибным чехлом, от которого отходят много численные гифы гриба. Эктотрофная микориза однолетняя. Она развивается летом или осенью и погибает к следующей весне, свойственна многим деревьям из семейств сосновых, буковых, березовых и др., а также некоторым травянистым растениям, например подъельнику. Эктотрофную микоризу чаще всего образуют базидиальные грибы из семейства Polyporaceae и особенно часто из рода Boletus. Так, подберезовик (В. scaber) образует микоризу на корнях березы, масленок – на корнях лиственницы (В. elegans) или сосны и ели (В. luteus), подосиновик (В. versipellis) – на корнях осины, белый гриб (В. edulus) – на корнях ели, дуба, березы (различные подвиды) и т. д. При эндотрофной микоризе мицелий гриба проникает в клетки корневой паренхимы корня, которые сохраняют свою жизнедеятельность, широко распространена у растений семейств орхидных, вересковых, брусничных, а также у многолетних трав из семейства сложноцветных и у некоторых деревьев, на пример у красного клена (Acer rubrum) и др. В качестве второго компонента эндотрофной микоризы часто выступает гриб Phoma из группы несовершенных грибов. Эндотрофная микориза может быть образована Oreomyces (живет на корнях орхидей, по–видимому, может фиксировать азот) и некоторыми другими видами грибов. Промежуточную форму микоризы, при которой имеет место и наружное обрастание корня гифами гриба, и проникновение гифов внутрь корня, называют перитрофной (эктоэндотрофной) микоризой. Значение микоризы в связи с ее широким распространением огромно. Многие растения – орхидные и вероятно вересковые, а также некоторые деревья без микоризы развиваются плохо или даже вообще не развиваются либо из–за недостатка питательных веществ в их мелких семенах, либо из–за недостаточного развития сосущих частей корешков, а также на бедных минеральными питательными веществами почвах. Грибы, образующие эндотрофную микоризу на их корнях, могут существовать лишь в кислой среде. Именно благодаря им многие представители орхидных и вересковых потому и живут только на кислых почвах. Следовательно, наличие в фитоценозе грибов, образующих микоризу, в значительной степени определяет видовой состав высших растений, входящих в этот фитоценоз, и служит немаловажным фактором в их борьбе за существование между растениями, поскольку отсутствие микоризы у растений, склонных к микотрофномy питанию, замедляет темпы их развития и ухудшает их положение по отношению к более быстро развивающимся видам, использующим микоризу. Грань между симбиозом и иными типами отношений иногда весьма условна. Интересно использование своей кишечной микрофлоры зайцеобразными и некоторыми грызунами. У кроликов, зайцев, пищух обнаружено регулярное поедание собственных фекалий. Кролики производят два типа экскрементов: сухие и мягкие, покрытые слизистой оболочкой. Мягкие фекалии они слизывают прямо с ануса и проглатывают не разжевывая. Исследования показали, что такая копрофагия вполне естественна. Кролики, лишенные возможности потреблять мягкий кал, худеют или плохо прибавляют в массе и чаще подвержены различным заболеваниям. Мягкий кал кроликов – это почти неизмененное содержимое слепой кишки, обогащенное витаминами (преимущественно В12) и белковыми веществами. Слепая кишка зайцеобразных представляет собой бродильный чан для переработки клетчатки и насыщена симбиотическими микроорганизмами. В 1 г мягкого кала насчитывается до 10 млрд бактерий. Попадая вместе с фекалиями в желудок кролика, микроорганизмы полностью погибают под влиянием кислоты и перевариваются в желудке и длинном тонком кишечнике. Таким образом, у исключительно растительноядных зайцеобразных копрофагия – это способ получения незаменимых аминокислот. Протокооперация (сотрудничество) – простой тип симбиотических связей. При этой форме совместное существование выгодно для обоих видов, но не обязательно для них, то есть не является непременным условием выживания видов (популяций). Например, дерево и гриб, термиты и жгутиковые простейшие, взаимоотношения птиц и носорога. Птицы кормятся насекомыми–паразитами на коже носорога, а их взлет служит ему сигналом опасности. Копытень и муравьи (рис. 114). Копытень растет в широколиственных и хвойно–широколиственных лесах. Цветет в конце апреля – мае. Венчик цветка грязно–пурпурный. Опыляется копытень мухами, но возможно и самоопыление. Семена копытня имеют мясистый придаток – ариллус. Муравьи поедают его, разносят семена и способствуют распространению растения.

Рисунок 114 – Копытень европейский (Asarum europeum L.)
Отрицательные взаимодействия в биоценозе Конкуренция – это взаимоотношения, возникающие между видами со сходными экологическими требованиями. Конкуренция возникает при взаимодействии популяций со сходным экологическим спектром, при наличии общих доступных ресурсов. Существует множество типов конкуренции. Прямая (интерференционная) конкуренция возникает при непосредственном контакте конкурирующих организмов. Косвенная (эксплуатационная) конкуренция возникает в результате уменьшении объема доступного ресурса. В сообществах конкурентные отношения существуют одновременно между несколькими видами. Тогда возникает диффузная конкуренция, то есть, конкуренция, обусловленная влиянием на популяцию всех видов, входящих в состав сообщества. В лабораторных условиях (в смешанных популяциях разных видов инфузорий–туфелек) был доказан принцип конкурентного исключения (правило Гаузе, или закон Гаузе): два вида не могут занимать одну экологическую нишу – в результате конкуренции в данной нише сохраняется только один, наиболее конкурентоспособный вид. Популяции менее конкурентоспособных видов «уходят» от конкуренции, формируя новые системы связей с окружающей их средой, новые экологические ниши. Таким образом, конкуренция способствует формированию новых экологических связей, образованию новых экологических ниш и, в конечном итоге, повышает уровень биологического разнообразия в сообществах. Рост численности конкурирующих популяций описывается системой уравнений Лотки–Вольтерра. Решение этой системы уравнений позволяет найти численности обеих популяций, при которых они могут существовать длительное время. Внутривидовая конкуренция. При внутривидовой конкуренции между особями сохраняются взаимоотношения, при которых они в состоянии размножаться и обеспечивать передачу свойственных им наследственных свойств. Внутривидовая конкуренция проявляется в территориальном поведении, когда, например, животное защищает место своего гнездовья или известную площадь в его округе. Так, в период размножения птиц самец охраняет определенную территорию, на которую кроме своей самки не допускает ни одной особи своего вида. Такую же картину можно наблюдать и у многих рыб (например, колюшки). Проявлением внутривидовой конкуренции является существование у животных социальной иерархии, которая характеризуется появлением в популяции доминирующих и подчиненных особей. Конкуренция между особями одного вида из–за пищи по мере увеличения плотности популяции становится более острой. В некоторых случаях внутривидовая конкуренция может приводить к дифференциации вида, к распадению его на несколько популяций, занимающих разные территории. Так, у саванной овсянки один экологический подвид размещается на сухих холмах, другой – на прибрежных солончаках. Конкуренция нередко является причиной переселения части популяции особей из одного географического района в другой. В условиях обостренной конкуренции за пищу или воду проявляется каннибализм у животных – регулятор численности популяции, способствующий установлению соответствия числа особей кормовым ресурсам и в целом благоприятствующий выживанию популяции. Постоянный, или облигатный каннибализм возник в процессе эволюции как полезное для вида в целом приспособление. Так, после спаривания самки богомолов поедают самцов (рис. 115). У некоторых организмов под влиянием внутривидовой конкуренции за жизненное пространство сформировался интересный тип поведения – территориальность. Она свойственна многим птицам, некоторым рыбам, другим животным. У птиц территориальность проявляется следующим образом. В начале сезона размножения самец выявляет «свою» территорию (участок обитания) и защищает ее от вторжения самцов того же вида. Заметим, что громкие голоса самцов, которые мы слышим весной, сигнализируют лишь о «праве собственности» на приглянувшийся участок, а вовсе не ставят себе задачу развлекать самку, как это обычно считается. Самец, строго охраняющий свой участок, имеет больше шансов успешно спариться и построить гнездо, тогда как самец, не способный обеспечить себе территорию, участвовать в размножении не будет. Иногда в охране территории принимает участие и самка. На охраняемой территории нелегкие заботы о гнезде и молоди не будут нарушены присутствием других родительских пар.

Рисунок 115 – Пример каннибализма
Защита территории вовсе не обязательно сопровождается активной борьбой. Громкого пения и угрожающих поз обычно достаточно для того, чтобы прогнать конкурента. Однако если один из партнеров–родителей погибает, его быстро замещает птица из числа еще не обосновавшихся особей. Таким образом, территориальное поведение можно считать регулятором, не допускающим как перенаселения, так и недоселенности. Яркий пример внутривидовой конкуренции, который каждый мог видеть в лесу, – так называемое самоизреживание у растений. Начинается этот процесс с захвата территории: например, где–нибудь на открытом месте, недалеко от большой ели, дающей множество семян, появляется несколько десятков всходов – небольших елочек. Первая задача выполнена – популяция подросла и захватила территорию, необходимую ей для выживания. Таким образом, территориальность у растений происходит иначе, нежели у животных: участок занимает не отдельная особь, а их группа (часть популяции). Молодые деревца растут, попутно затеняя и угнетая находящиеся под их кронами травянистые растения (это уже пример межвидовой конкуренции). С течением времени между деревцами появляется неизбежная разница в росте: одни, что слабее, отстают, другие – обгоняют. Поскольку ель – очень светолюбивое растение (ее крона поглощает почти весь падающий свет), то более слабые молодые елочки начинают все больше испытывать затенение со стороны высоких деревьев и постепенно засыхают, отмирают. В конце концов, через много лет на поляне от сотни елочек остается всего два–три деревца (наиболее сильные особи из всего поколения). Но зато никаких трав уже нет, корни деревьев раскинулись по всей территории поляны, и ничто не мешает тянуть еще выше к солнцу пышную хвойную крону. У некоторых видов внутривидовое регулирование начинается задолго до того, как обнаруживается серьезная конкуренция. Так, высокая плотность животных является фактором угнетения, снижающим скорость воспроизводства этой популяции даже при изобилии пищевых ресурсов. Внутривидовая конкуренция является важным регулятором, контролирующим рост популяций. Благодаря этой конкуренции возникает определенная зависимость между плотностью популяции и скоростью процессов отмирания (смертность) или размножения (рождаемость) особей. Это, в свою очередь, приводит к возникновению определенной связи между численностью родительских пар и количеством производимого ими потомства. Подобные связи действуют как регуляторы колебаний численности популяций. Межвидовой конкуренцией называют активный поиск двумя или несколькими видами одних и тех же пищевых ресурсов среды обитания. Конкурентные взаимоотношения, как правило, возникают между видами со сходными экологическими требованиями. При совместном обитании каждый из них находится в невыгодном положении в связи с тем, что присутствие другого вида уменьшает возможности в овладении пищевыми ресурсами, убежищами и другими средствами к существованию, имеющимися в местообитании. Конкуренция относится к форме экологических отношений, отрицательно сказывающейся на взаимодействующих партнерах. Конкурентные взаимоотношения могут быть самыми различными – от прямой физической борьбы до мирного совместного существования. Например, на Крайнем Севере Дальнего Востока береза каменная, ольховый и кедровый стланики формируют чистые сообщества и сообщества с доминированием одного из них на склонах южных экспозиций. Часто они произрастают совместно и доминанта трудно выделить. Все три вида характеризуются очень близкими экологическими свойствами. Все они – реликтовые, и отличаются высоким тепло–, влаго– и светолюбием. Но при этом ольховник несколько более теневынослив и более требователен к влажности почв, береза – более требовательна к теплу и трофности почв, а кедровый стланик – к свету и влажности воздуха. В результате при совместном произрастании кедровостланиковые ценоэлементы, или парцеллы, обычно приурочены к повышенным элементам микрорельефа, более сухим и хорошо дренированным, каменноберезовые – к выровненным участкам с высокой трофностью и тоже хорошо дренированных, ольховниковые – к микропонижениям с очень высоким, но проточным увлажнением и высокой трофностью почв. Каменноберезняки чаще приурочены к распадкам и в горах не поднимаются выше стлаников, кедровый стланик образует чистые заросли на верхней границе леса и на грядах, расположенных полосами вдоль по склону, а заросли ольховника предпочитают седловины и перегибы поверхностей склона в местах с вогнутой поверхностью. Таким образом, в каждом фитоценозе подбираются растения: представляющие различные жизненные формы и занимающие место в различных синузиях, ярусах, микроценозах, то есть образующие группы, характеризующиеся неодинаковым отношением к среде и неодинаковым местом в фитоценозе; дифференцированные по срокам прохождения сезонных фаз. Сочетание в одном фитоценозе растений с различными экологическими особенностями – тенелюбивых и светолюбивых, в различной степени приспособленных к недостатку влаги и другим факторам среды, позволяет фитоценозу наиболее полно использовать условия местообитания. Вытеснение одних видов другими в фитоценозе может происходить в результате различной конкурентной способности видов, в частности различной энергии вегетативного размножения. Например, всем известный сорный злак пырей не только способен заглушить культурные посевы, но и вытесняет многие дикие виды (крапиву, чистотел и др.), растущие с ним по соседству и которые вегетативно размножаются очень слабо. Даже клевер ползучий постепенно уступает позиции пырею. Очень сильной конкурентной способностью обладает мох сфагнум. По мере разрастания он в прямом смысле поглощает соседние растения. В районах распространения вечной мерзлоты фитоценозы с господством сфагнума занимают обширные пространства, вытесняя зоны своего влияния не только травы и кустарнички, но и кустарники и деревья. В результате борьбы за существование происходит дифференциация видов, образующих фитоценоз. В то же время строение фитоценоза – не только результат борьбы за существование, но и результат приспособления растений к уменьшению напряженности этой борьбы. В фитоценозе виды подобраны таким образом, что дополняют друг друга своими свойствами. Интересным примером межвидовой конкуренции у растений и бактерий является аллелопатия. Оказалось, что некоторые организмы способны выделять в окружающую среду (например, в почву или в воздух) химические вещества (алколоиды, терпены), подавляющие развитие конкурентов. Например, гриб–пеницилл продуцирует вещества, угнетающие жизнедеятельность бактерий. Вещества, выделяемые надземными и подземными органами живых растений, и органические соединения, получающиеся в процессе разложения мертвых растительных остатков и оказывающие влияние на другие растения, называются колинами. Колины образуются как при свободной жизни растений рядом друг с другом, так и при паразитизме и взаимном влиянии пыльцы разных видов. Выделения веществ могут оказывать положительное или отрицательное влияние на растения того же вида или других видов. В частности, антибиотические вещества, препятствующие росту различных плесеней и бактерий, выделяют плесневые грибы, бактерии, актиномицеты, а также некоторые водоросли и лишайники. Эти выделения защищают растения от нападающих на них паразитов и тем самым ограничивают число видов, входящих в состав фитоценоза, или затрудняют развитие некоторых из них. Среди колинов различают: – газообразные выделения надземных органов растений, – прочие выделения наземных органов растений, – корневые выделения, – продукты распада мертвых растительных остатков. Среди газообразных выделений большую роль играет этилен, в значительных количествах вырабатываемый некоторыми растениями, например, яблоками. (Этилен задерживает рост, вызывает преждевременный опад листвы, ускоряет распускание почек и созревание плодов, оказывает положительное или отрицательное влияние на рост корней). Газообразные колины могут оказать воздействие на ход сезонных явлений в ценозе, а также подавить развитие тех или иных видов. Однако более или менее значительной роль газообразных колинов может быть лишь в аридных регионах, где обильно число растений, выделяющих различные легко испаряющиеся эфирные масла. Эти эфирные масла служат приспособлением к понижению температуры вокруг испаряющей поверхности, но одновременно могут оказывать определенное воздействие на те или иные растения. Твердые и жидкие выделения надземных органов растений представляют собой минеральные и сложные органические соединения, которые вымываются из надземных частей растений осадками, иногда в весьма значительных количествах, и оказывают действие на другие растения, попадая на них непосредственно с дождем, росой или через почву. Так, выделения полыни горькой (Artemisia absinthium) задерживают рост многих растений, то же указывается для веществ, содержащихся в листьях ореха черного (Juglans nigra), а также в листьях и хвое многих древесных пород и некоторых кустарников и трав. Нейтрализм – оба вида независимы и не оказывают друг на друга никакого влияния. При нейтрализме особи не связаны друг с другом непосредственно, и сожительство их на одной территории не влечет для них как положительных, так и отрицательных последствий, это зависит от состояния сообщества в целом. Так, лоси и белки, синицы и мыши, обитающие в одном лесу, практически не контактируют друг с другом. Аменсализм – тип межвидовых взаимоотношений, при котором в совместной среде обитания один вид подавляет существование другого вида, не испытывая противодействия (например, у растений, светолюбивые виды, растущие под елью, испытывают угнетение в результате сильного затенения кроной, тогда как для самого дерева их соседство может быть безразличным). Хищничество – такой тип взаимоотношений, при котором представители одного вида поедают представителей другого, то есть организмы одного вида служат пищей для другого. Например, взаимоотношения волков и северных оленей (рис. 116). Одним из немногих примеров хищничества в мире растений являются росянки (рис. 117). Испытывая азотистое голодание на бедных почвах болот, росянка получает необходимые вещества, отлавливая и переваривая мелких беспозвоночных. Ловчим аппаратом росянке служат листья; на каждом листе насчитывается до 200 чувствительных волосков. Также на листьях расположены железы, выделяющие липкий секрет. Для перевода сложных органических соединений в доступную для растений форму, росянка использует специфический ферментативный аппарат, близкий по составу к животному. Различают внутривидовую и межвидовую конкуренцию. Чем в более родственных отношениях находятся популяции, тем более жесткими являются отношения между ними, т.к. они нуждаются в сходных экологических ресурсах, которых, как правило, не хватает.

Рисунок 116 – Отношения хищник–жертва (волки и северные олени)
Хищник, как правило, вначале ловит жертву, убивает ее, а за тем поедает. Для этого у него имеются специальные приспособления. У жертв также исторически выработались защитные свойства в виде анатомо–морфологических, физиологических, биохимических особенностей. Например, выросты тела, шипы, колючки, панцири, защитная окраска, ядовитые железы, способность быстро прятаться, зарываться в рыхлый грунт, строить недоступные хищникам убежища, прибегать к сигнализации об опасности. Вследствие таких обоюдных приспособлений формируются определенные группировки организмов в виде специализированных хищников и специализированных жертв. Так, основной пищей рыси служат зайцы, а волк типичный многоядный хищник.

Рисунок 117 – Росянка круглолистная (Drosera rotundifolia L.)
Большинство плотоядных животных, как было отмечено, в состоянии убить и сразу же съесть свою жертву, так как превосходят ее размерами и силой. Популяции, находящиеся на разных трофических уровнях, могут вступать во взаимоотношения хищник–жертва (рис. 118). Во многих случаях циклические колебания численности популяции хищника сдвинуты по фазе по отношению к колебаниям численности популяции жертвы. Это можно объяснить тем, что возрастание численности хищника приводит к резкому снижению численности жертвы, вслед за чем в результате голода уменьшается количество хищников. Это позволяет снова вырасти числу жертв, вслед за чем начинает расти количество хищников. Характерный период таких циклов обычно составляет несколько поколений.
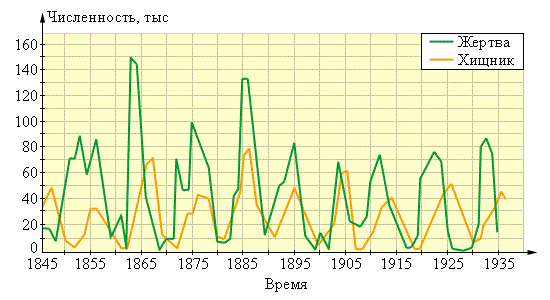
Рисунок 118 – Колебания численности популяций хищника и его жертвы
Некоторые организмы несъедобны для их потенциальных хищников (так, ряд бабочек ядовиты для птиц). Часто эти организмы имеют яркую предупреждающую окраску; хищники избегают охотиться на окрашенных таким образом жертв. В качестве примера предупреждающей окраски можно привести некоторых бабочек и жуков. Этим пользуются другие, уже неядовитые организмы, подражающие своей окраской ядовитым видам. Другие животные подражают различным объектам растительного или животного мира или даже каким–либо минералам. Подражание другим объектам с целью защиты от врагов (мимикрия) широко распространённое среди насекомых, встречается и среди многих других групп животных (например, у некоторых неядовитых змей) (рис. 119).

Рисунок 119 – Пример защиты от врагов – мимикрия
Слева – гусеница, имитирующая сучок растения Справа – бабочка вице–король повторяет форму и окраску крыльев ядовитой бабочки–монарха
Как правило, численность моделей особей должна быть значительно выше, чем численность подражателей, главным образом для того, чтобы хищники быстро научились избегать ядовитых форм. Существует также кольцо мимикрии, когда несколько несъедобных для хищников видов имеют схожую окраску (например, различные виды ос) с тем, чтобы враги, выработав соответствующий рефлекс на один вид, не трогали и особей других видов. Результатом конкуренции может быть полное вытеснение одной из популяций из экосистемы или сужение ее условий существования до границ, определяемых жизнеспособностью доминирующей популяции. Так, известно, что серая крыса вытеснила черную, будучи более крупной, более агрессивной, способной преодолевать различные препятствия и адаптироваться к изменяющимся условиям существования человека. В Австралии завозная пчела вытеснила туземную, маленькую, без жала. Особой формой конкурентных отношений между популяциями за ресурсы среды (в первую очередь за пищевые ресурсы), приводящей к подавлению репродукции одной из популяций к ее уничтожению, является интерференция. Этот термин изначально предполагалось использовать вместо не очень точного термина «конкуренция», всегда требующего уточнения вида конкуренции. В биологической литературе последних лет издания под интерференцией рассматривают «взаимодействия (обычно неблагоприятные), возникающие при наличии у одних организмов близких соседей того же или других видов. Примером интерференции может быть попутное уничтожение зимующих в шишках ели насекомых клестами в процессе питания или мелких насекомых крупными при питании одним и тем же субстратом» (Г.В. Стадницкий и др.,1997, с.114). В « Биологическом энциклопедическом словаре» (с.232) представлена интерференция вирусов – «тип взаимодействия между вирусами, при котором наблюдается подавление репродукции одного вируса другим в клетках, смешанно зараженных вирусами». Паразитизм – это форма взаимоотношений между видами, при которой организмы одного вида (паразиты, потребители) живут за счет питательных веществ или тканей организма другого вида (хозяина) в течение определенного времени. Обычно паразит использует живого хозяина не только как источник пищи, но и как место постоянного или временного проживания. В роли паразита могут выступать и растения, и животные. В отличие от хищничества при нападении паразита хозяин не погибает сразу, но испытывает угнетение (нередко в течение длительного времени). Другими словами, паразит изнуряет, но не губит хозяина, поскольку это обеспечивает его существование. Таким образом, паразитизм можно рассматривать как ослабленную форму хищничества. Паразитизм в отличие от хищничества характеризуется тремя основными особенностями. Паразит в течение всей своей жизни нападает всего на одну особь (редко – на многих) и поедает только часть вещества своей жертвы (хозяина); паразит причиняет хозяину вред, но очень редко приводит к его быстрой гибели; Паразит обязательно живет (постоянно или временно) в теле или на поверхности тела своего хозяина, поэтому паразиты обычно намного мельче хозяев; Паразит гораздо теснее связан со своим хозяином, чем хищник с жертвой. Это результат естественного отбора и узкой специализации видов. Ленточные черви, печеночная двуустка, вирус кори, туберкулезная палочка – все это обычные примеры паразитов, которые поражают животных. Можно привести длинный список паразитов, вредящих растениям. К ним нередко относятся сами же растения, грибы и микроорганизмы. Таковы, например, повилика, заразиха, фитофтора, вирус табачной мозаики, головневые и ржавчинные грибы. Некоторых насекомых выделяют в отдельную экологическую группу паразитоидов. Взрослые насекомые этой группы ведут свободный образ жизни, но яйца откладывают либо в тело личинки другого насекомого, либо на поверхность его, а иногда – в тело пауков и мокриц. Вылупившиеся из яиц личинки развиваются в теле своего хозяина, питаясь его тканями. Вначале личинка наносит хозяину незначительный вред, но по мере своего развития она почти целиком съедает его. В конце концов, из того, что было когда–то куколкой или личинкой хозяина, появляется взрослое насекомое–паразитоид. Паразитоиды занимают промежуточное положение между хищниками и паразитами. С хищниками их сближает то, что от их действий хозяин погибает. Однако настоящий хищник непосредственно нападает на свою жертву и питается ею. Нападением паразитоида на жертву (хозяина) является откладывание самкой яйца, хотя в действительности именно личинка, которая вывелась из этого яйца питается за счет хозяина. Экологические категории паразитов. Истинный паразитизм характеризуется узкой специализацией видов, поскольку хозяин обеспечивает паразиту и пищу, и микроклимат, и защиту, и местообитание. Чем лучше паразит приспособлен к особенностям организма хозяина, тем успешнее его размножение и развитие потомства. Каждый год миллионы людей гибнут от различных инфекций, становятся инвалидами или калеками в результате заражения паразитами. Люди, образуя плотные и скученные популяции и создавая такие же условия для домашних животных и культурных растений, сами способствуют крайне благоприятной обстановке для паразитарных инфекций. Даже в дикой природе лишь редкие из свободноживущих видов организмов не поражены хотя бы немногими особями паразитических видов. Паразитов подразделяют на две основные категории: микропаразитов и макропаразитов. К микропаразитам относятся те, которые непосредственно размножаются внутри тела хозяина, обычно внутри клеток. Макропаразиты растут в теле хозяина, но размножаются, образуя особые формы, которые покидают хозяина, чтобы заселить нового. Наиболее известными микропаразитами являются бактерии и вирусы, например возбудители кори и тифа, а также листовой мозаики культурных растений. Другая важнейшая группа микропаразитов, поражающих животных, – это представители простейших, например возбудитель малярии. Микропаразитов можно разделить на тех, которые передаются от одного хозяина к другому непосредственно, и тех, которые передаются при участии другого вида – переносчика. Непосредственная передача возбудителя болезни бывает почти мгновенной, как это происходит при передаче паразитов, способных короткое время существовать в образующихся при кашле или чихании капельках влаги (грипп, корь и т. д.). В других случаях паразит продолжительное время находится в покоящейся стадии, «поджидая» нового хозяина, и попадает в него при поглощении им пищи или воды (например, из простейших амеба, вызывающая дизентерию). Обычное местообитание макропаразитов – различные полости тела. Один из главных макропаразитов животных – глисты, или гельминты, к которым относятся ленточные черви (из типа плоских червей), ряд видов круглых червей. Кроме того, животных поражают вши, блохи, клещи, а также грибы. К макропаразитам растений относятся грибы, вызывающие мучнистую росу, ржавчину, головню и другие виды заболеваний, а также цветковые растения – повилика, заразиха, раффлезия. Циклы развития и передача паразитов. Некоторые паразиты человека имеют очень сложный жизненный цикл. Ленточные черви, например, во взрослом состоянии живут в кишечнике, поглощая питательные вещества хозяина непосредственно поверхностью своего тела, которое состоит из большого числа постоянно нарастающих члеников. Самые последние из этих члеников, содержащее в себе многочисленные яйца, постепенно отпадают от тела червя и выводятся из организма хозяина вместе с экскрементами. Для нормального развития личинок яйца должны попасть в организм промежуточного хозяина – того животного, в теле которого проходит одна из стадий развития паразита. После этого паразит вновь возвращается (чаще всего с пищей) в организм основного хозяина (человека), где происходит его размножение. Для некоторых видов ленточных червей человек может быть не основным, а промежуточным хозяином, как, например, при заражении эхинококком. Личинки эхинококка, содержащиеся в плохо обработанной свинине, проникнув в ткани различных внутренних органов человека, особенно печени, легких, развиваются до величины огромных пузырей. Передача паразита от одного хозяина к другому является важнейшей стадией любого паразитного заболевания. От особенностей передачи во многом зависит скорость распространения возбудителей болезни в популяции хозяина. Когда макропаразиты передаются непосредственно, заражение происходит при физическом контакте между особями хозяина и скорость распространения возбудителя обычно прямо пропорциональна частоте встреч зараженных хозяев с восприимчивыми незараженными. Следовательно, в плотной популяции хозяина скорость всегда выше, нежели в разряженной. Важное значение при этом имеют климатические факторы, оказывающие влияние на устойчивость короткоживущих инфекционных стадий, а также сезонные изменения частоты контактов, играющие важную роль в эпидемиологии – науке, изучающей факторы и характер распространения эпидемий многих заболеваний человека (кори, свинки, ветряной оспы). Иммунный ответ. Огромное значение в динамике распространения инфекционных болезней имеет иммунный ответ, то есть способность организма хозяина к выздоровлению после заражения, а также образующаяся в ходе борьбы с болезнью «память» организма, способная обеспечить его невосприимчивость к данной инфекции – иммунитет. У млекопитающих, в том числе и у человека, иммунитет к некоторым видам инфекций может передаваться даже потомкам. Большинство вирусных и бактериальных заболеваний высших животных – лишь кратковременный эпизод их жизни. Паразиты размножаются в теле хозяина, вызывая мощный иммунный ответ. И напротив, такой ответ на многих макропаразитов и простейших, как правило, довольно непродолжителен. Поэтому заселение ими хозяина обычно устойчиво, причем хозяева подвергаются непрерывному перезаражению. Популяционная динамика паразитизма. Между динамикой взаимодействия паразита – хозяина и хищника – жертвы много общего. Например, повторяющиеся (циклические) вспышки эпидемии кори и других детских болезней в точности соответствуют колебаниям численности хищника и жертвы. Это сходство определяется тем, что приобретение иммунитета переболевшими особями влияет на популяцию паразитов так же, как снижение плотности популяции жертв на популяцию хищника. В некоторых случаях, однако, динамика паразитизма может сильно отличаться от динамики хищничества в силу того, что популяция паразита зависит не только от популяции основного хозяина, но и от популяций промежуточных хозяев и переносчиков. Периодические вспышки заболеваемости, характерные для многих инфекций, в том числе для гриппа и кори, отражают последовательное чередование «высокой частоты заболеваемости», «малой плотности восприимчивых особей», «низкой частоты заболеваемости», «высокой плотности восприимчивых хозяев», снова «высокой частоты заболеваемости» и т. д. Приобретение иммунитета уменьшает число восприимчивых организмов. Полный иммунитет означает, что это число становится равным нулю. Таким образом, типичная картина эпидемии такова: вначале вся популяция восприимчива к инфекции и заболевание распространяется очень быстро; иммунитет отсутствует, и каждый заболевший передает инфекцию всем, с кем он вступал в контакт; однако в дальнейшем скорость распространения заболевания начинает снижаться, так как у заболевших вырабатывается иммунитет. К концу эпидемии популяция либо полностью приобретает иммунитет, либо вымирает. Восприимчивых к инфекции фактически не остается. Различают следующие виды паразитизма: а) экзопаразитизм и эндопаразитизм – в первом случае паразит находится на поверхности тела хозяина, во втором случае – внутри тела хозяина; б) временный и стационарный – в первом случае паразит нападает на хозяина только для питания, во втором случае – проводит на хозяине большую часть жизни. в) факультативный и облигатный. Первые могут вести и паразитический, и сапрофитный образ жизни, как, например, грибы трутовики из рода полипорус (Polyporus), вторые ведут только паразитический образ жизни. В связи с этим облигатные паразиты обычно не сразу губят своего хозяина, а постепенно истощают его, в то время как факультативные паразиты весьма быстро приводят своего хозяина к гибели и переходят на питание его мертвыми тканями. Поэтому факультативные паразиты оказывают более существенное воздействие на растение–хозяина, чем облигатные. К числу паразитов относятся многие грибы, некоторые слизевики, водоросли и лишайники, а также ряд видов цветковых растений. Так, повилика европейская (Cuscuta europaea) иногда сильно поражает крапиву двудомную (Urtica dioica), а повилика хмелевидная (С. lupuliformis) и однопестичная (С. Monogyna) – иву (Salix). Паразитами следует считать и бактерии, вызывающие заболевания растений, животных и человека. Кроме полных паразитов, существуют гемипаразиты (полупаразиты). Последние, имея хлорофилл, способны к ассимиляции и от растения–хозяина получают преимущественно минеральное питание в водном растворе. Часть полупаразитов представляют виды, обычно обитающие на лугах, иногда в лесах, с очень слабыми корневыми системами. Эти растения присасываются своими корнями к корням цветковых растений. Это многие виды семейства норичниковых (Scrophulariaceae) из родов мытник (Pedicularis), марьянник (Melampyrum), зубянка (Odontites), очанка (Euphrasia), погремок (Alectorolophus) и др. Массовое их развитие может резко снизить качество луга, так как ценные в хозяйственном отношении травы сильно истощаются и резко снижают урожай. Различают полный паразитизм и полупаразитизм. Например, повилика – это вьющееся растение–паразит не имеет листьев (они редуцированы до мелких чешуек), корней, красноватый стебель нитевидный, обвивающий различные растения. Паразитирует в основном на крапиве, также на хмеле, клевере, горошке и других растениях. На стебле повилики образуются присоски (гаустории), через которые из растения–хозяина повилика добывает питательные вещества (рис. 120). Примером растения–полупаразита может служить растение погремок. Обладая зелеными фотосинтезирующими листьями и слабо развитыми корнями, он «присасывается» к корневой системе растущих по соседству злаков и частично живет за их счет (рис. 121). В условиях умеренного климата наиболее широко распространена омела белая (Viscum album) на лиственных, реже на хвойных породах. При паразитизме во многих случаях наблюдается смена хозяев. Разным стадиям жизненного цикла паразитов часто соответствуют разные (обычно строго определенные) хозяева. Численность популяций при взаимодействиях «паразит–хозяин» также регулируется системой обратных связей. Но в отличие от хищников паразиты любого типа оказывают меньшее влияние на численность эксплуатируемой популяции, но зато сами паразиты вследствие специализации гораздо сильнее зависят от эксплуатируемой популяции, чем хищник от жертвы. Поэтому амплитуда колебаний численности паразита более тесно связана с амплитудой колебаний численности хозяина, но фазовый сдвиг значительно меньше, чем в системе «хищник–жертва».

Рисунок 120 – Повилика (Cuscuta L.)

Рисунок 121 – Погремок узколистный (Rhinanthus angustifolius C. C. Gmel.)
И хищник, и паразит могут существовать за счет одного или нескольких видов. При этом различают следующие типы: полифаги – нападающие на большое число видов. К ним относятся многие хищные млекопитающие и насекомые. Насекомые–фитофаги питаются самыми различными растениями. Так, гусеница стеблевого мотылька поражает более чем 200 видов растений; олигофаги – живущие за счет нескольких, часто близких видов. Колорадский жук питается картофелем и другими растениями, относящимися главным образом к пасленовым. Солитер паразитирует в человеке, свинье, различных плотоядных и грызунах; монофаги – живущие за счет только одного хозяина. Монофагия является правилом для многих паразитических насекомых, например, таких, как грушевый цветоед – на груше, шелковичный червь – на тутовом дереве. Переход к паразитическому образу жизни обычно сопровождается дегенерацией. Утрачиваются органы пищеварения, органы чувств, теряется подвижность. У паразитов изменяется форма тела, развиваются разнообразные крючья и присоски. Жизненный цикл паразитов сопряжён с огромной смертностью; следствием этого является повышенная способность к размножению (иногда измеряемая в миллионах особей в год). К паразитам относятся многие болезнетворные бактерии и простейшие, некоторые грибы, вьющиеся лианы; паразитические формы имеются среди представителей большинства типов животных (рис. 122).
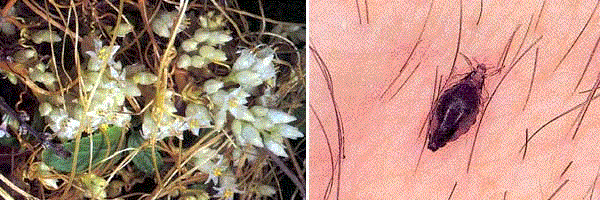
Рисунок 122 – Паразитические формы сосуществования. Слева направо: вьющиеся лианы, платяная вошь
Душители – корнесобственные растения, но начинают развитие в качестве эпифитов. Различные животные переносят их семена с одного дерева на другое. Птицы – основные переносчики семян душителей. Душитель образует корни двух родов: одни из них плотно прилегают к коре дерева–хозяина, ветвятся, и образуют плотную сеть, одевающую ствол дерева–хозяина, другие свешиваются вертикально вниз и, достигнув почвы, ветвятся в ней, доставляя душителю воду и минеральное питание. В результате затенения и сдавливания дерево–хозяин погибает, а душитель, к тому времени развивший мощный корневой «ствол», остается стоять на «собственных ногах». Многочисленные лианы фестонами свисают с дерева. Душители характерны для влажных тропиков. Душители находятся с деревьями–хозяевами в антагонистических отношениях. Некоторые южноамериканские виды–душители имеют настолько слабые корни, что при падении дерево–хозяин увлекает их за собой. Наибольшую опасность представляют паразиты для ослабленных особей и популяций в естественных экосистемах, а также для всех организмов в антропогенных экосистемах, вследствие их гомогенности и низкой пластичности. Форма паразитизма, при которой одна популяция использует другую для обеспечения собственной жизнедеятельности, называется рабовладельчеством. Так, муравьи–амазонки (Polyergus rufescens) похищают личинок и куколок из других муравейников, выводят из них взрослых насекомых, которые выполняют работы по уходу за потомством хозяев, а также по уходу и достройке жилища. Еще одна любопытная форма паразитизма – клептопаразитизм, присвоение корма добытого особями одной популяции особями другой. Он часто наблюдается у многих видов птиц, рыб и млекопитающих. Так короткохвостый поморник обеспечивает себя и свое потомство кормом (рыбой), которую он отбирает у типиков, маевок и кайр. Чайки также часто преследуют и нападают на топорков, крачек и других птиц, заставляя их бросать корм, который подхватывают на лету. Все перечисленные выше типы взаимоотношений и их общие характеристики сведены в таблице.
Таблица 11 – Типы биотических взаимоотношений
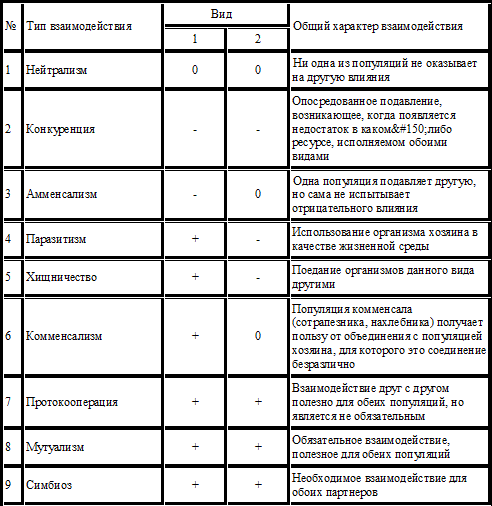
Следует помнить, что типы взаимоотношений конкретной пары видов могут изменяться в зависимости от внешних условий или стадии жизни взаимодействующих организмов. К тому же в природе во взаимоотношения оказывается вовлеченной не пара видов, а гораздо большее число. Межвидовые связи в природе бесконечно разнообразны.
|