Тема 3.3. Пространственная структура сообществ
Пространственная структура наземного биоценоза определяется прежде всего сложением его растительной части – фитоценоза, распределением наземной и подземной массы растений. Пространственная структура растительного сообщества включает: надземную и подземную ярусность, синузиальность и мозаичность фитоценоза. Ярусностью фитоценоза называют размещение органов растений различных видов на разных высотах над поверхностью почвы и на разных глубинах в почве.Различают ярусность надземную и подземную. Распределение растений по надземным ярусам определяет неодинаковая освещенность, которая приводит к различиям в температурном режиме и режиме влажности.
В одних и тех же ярусах находятся растения одинаковой высоты, сходные или различающиеся по своим экологическим особенностям (например, хвойные и лиственные породы), но имеющие примерно одинаковую потребность в освещении. Растения разных ярусов влияют друг на друга. Растения верхних надземных ярусов более светолюбивы, чем растения нижних ярусов, и лучше приспособлены к колебаниям температуры и влажности. Под своими кронами они создают условия слабой освещенности и стабильной температуры и влажности. Поэтому нижние ярусы образованы растениями, у которых потребность в свете меньше. В свою очередь растения нижних ярусов влияют на растения верхних ярусов. Так, например, ярус мхов в еловом или пихтовом лесу накапливает значительное количество влаги; травяной покров леса участвует в процессе почвообразования, формируя опад, и т.д. К тому или иному ярусу следует относить растения, достигшие обычных для данного вида размеров. Молодые же растения, временно входящие в состав более низких ярусов не должны в них включаться. Не следует выделять в особый ярус те экземпляры того или иного вида, которые временно настолько угнетены, что не способны размножаться семенным или вегетативным путем. Так, если в результате выборочной рубки в смешанном липово–еловом лесу, где липа образовывала второй ярус под покровом ели, липа приняла кустарниковую форму и перестала размножаться как семенным, так и вегетативным путем, ее нельзя выделять в особый ярус. В этом случае причиной того, что липа приняла форму кустарника, является постоянная рубка более высоких стволов этого дерева. Если рубку прекратить, то липа принимает свойственную ей древесную форму. Если же липа в лесу под влиянием затенения приобретает кустарниковую форму и теряет способность к семенному размножению, хотя обычно при этом размножается вегетативно, ее следует выделить в особый ярус. В тех случаях, когда угнетенное состояние растения, обитающего в одном из нижних ярусов, длится долго, их нужно причислять к этому ярусу и при отсутствии способности к вегетативному размножению. Молодые растения постепенно вырастают. Поэтому их, так же как растения, низкорослость которых поддерживается каким–либо посторонним по отношению к фитоценозу фактором, относят к пологу. Пологом (Сукачев, 1930) называют временный ярус, образованный молодыми растениями или растениями, угнетенными посторонними по отношению к фитоценозу факторами (например, рубкой). Таким образом, каждый вид входит в состав только одного яруса, а в других ярусах экземпляры этого вида присутствуют временно и временно образуют полога, располагающиеся между ярусами фитоценоза. Ярусность особенно хорошо заметна в лесах умеренного пояса. Например, в еловых лесах четко выделяются древесный, травяно–кустарничковый и моховый ярусы. Пять или шесть ярусов можно выделить и в широколиственном лесу: первый, или верхний, ярус образован деревьями первой величины (дуб черешчатый, липа сердцевидная, клен платановидный, вяз гладкий и др.); второй – деревьями второй величины (рябина обыкновенная, дикие яблони и груша, черемуха, ива козья и др.); третий ярус составляет подлесок, образованный кустарниками (лещина обыкновенная, крушина ломкая, жимолость лесная, бересклет европейский и др.); четвертый состоит из высоких трав (борцы, бор развесистый, чистец лесной и др.); пятый ярус сложен из трав более низких (сныть обыкновенная, осока волосистая, пролесник многолетний и др.); в шестом ярусе – наиболее низкие травы, такие, как копытень европейский. Подрост деревьев и кустарников может быть разного возраста и разной величины и не образует особых ярусов. Наиболее многоярусны дождевые тропические леса, наименее – искусственные лесные насаждения (рис. 123, 124). В лесах всегда есть и межъярусные (внеярусные) растения – это водоросли и лишайники на стволах и ветвях деревьев, высшие споровые и цветковые эпифиты, лианы и др.

Рисунок 123 – Многоярусный дождевой тропический лес Центральной Амазонки. Растительность полосы длиной 20 м и шириной 5 м
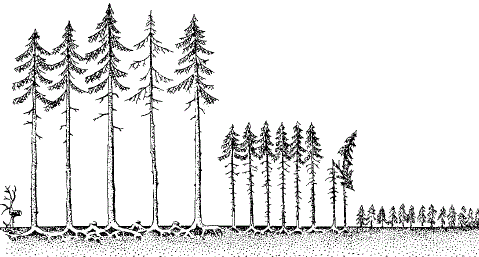
Рисунок 124 – Одноярусный саженый еловый лес. Монокультуры разного возраста
Ярусность позволяет растениям более полно использовать световой поток: под пологом высоких растений могут существовать теневыносливые, вплоть до тенелюбивых, перехватывая даже слабый солнечный свет. Ярусность выражена и в травянистых сообществах (лугах, степях, саваннах), но не всегда достаточно отчетливо (рис. 125). Кроме того, в них обычно выделяют меньше ярусов, чем в лесах. Впрочем, и в лесах иногда насчитываются всего два четко выраженных яруса, например в бору–беломошнике (древесный, образованный сосной, и напочвенный – из лишайников).

Рисунок 125 – Ярусность растительности луговой степи (по В. В. Алехину, А. А. Уранову, 1933)
Ярусы выделяют по основной массе ассимилирующих органов растений, оказывающих большое влияние на среду. Ярусы растительности могут быть разной протяженности: древесный ярус, например, толщиной в несколько метров, а моховой покров – всего несколько сантиметров. Каждый ярус по–своему участвует в создании фитоклимата и приспособлен к определенному комплексу условий. Например, в еловом лесу растения травяно–кустарничкового яруса (кислица обыкновенная, майник двулистный, черника и др.) находятся в условиях ослабленного освещения, выровненных температур (более низких днем и более высоких ночью), слабого ветра, повышенных влажности и содержания СО2. Таким образом, древесный и травяно–кустарничковый ярусы находятся в разной экологической обстановке, что сказывается на функционировании растений и на жизни животных, обитающих в пределах этих ярусов. Не все ярусы одинаковы. Одни из них, образованные только деревьями или только кустарниками и кустарничками, постоянны и сохраняют систему стволов и ветвей, а в ряде случаев и листьев, круглогодично. Другие же непостоянны. Они образованы травянистыми растениями, надземные части которых полностью или частично отмирают на неблагоприятный период года. Говорить об оформленном ярусе можно только в том случае, когда растения в данном ярусе стоят достаточно густо, в результате чего затенение заставляет их вытягиваться до одного общего уровня. Взаимное влияние растений, образующих ярус, выражается степенью проективного покрытия этого яруса. О невыраженной ярусности, то есть о разновысотности растений говорят, когда такой сомкнутости нет (Раменский, 1938). Так, на рисунке 126 верхний (первый) ярус тропического леса не выражен, а второй и третий выражены, но не разобщены один от другого, и провести границу между ними очень трудно. Степень выраженности ярусов хорошо заметна на диаграммах, показывающих степень проективного покрытия фитоценоза по ярусам. На рисунке 127 показано, что в дубовом лесу (Чехия) ярус кустарников из волчьего лыка и ярус напочвенного мохового покрова, выражены плохо. А древесные ярусы 1 и 2 и ярус травянистых растений выражены хорошо.

Рисунок 126 – Вертикальная проекция первичного смешанного тропического леса
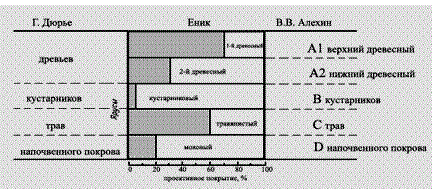
Рисунок 127 – Ярусность дубравы в лесничестве Држевич (по Енику) и в понимании Г. Дюрье и В.В. Алехина
При выделении ярусов выделяют два (или три) яруса деревьев, один или два яруса кустарников, три яруса трав, один ярус напочвенного покрова. В. В. Алехин (1950) предлагал древесным ярусам присвоить буквенный индекс А и называть A1 – верхний древесный ярус, А2 – средний древесный ярус и т. д., ярус кустарников он обозначил индексом В, ярус трав – С, ярус напочвенного покрова – D. Г. Дюрье выделяет крупные ярусы: деревьев, кустарников, трав, напочвенного покрова, а в пределах этих ярусов – подъярусы. В. Н. Сукачев намечает ярусы: первый, второй, если нужно – третий древесный, кустарников, трав, напочвенного покрова. При этом последние три яруса (кустарников, трав и напочвенного покрова) можно подразделять на подъярусы. Эпифиты и лианы очень часто развивают свою листву в определенных ярусах леса. Так, крупные лианы обычно имеют длинные безлистные стебли, и их крона развивается в кронах деревьев того или иного яруса. Другие виды эпифитов и лиан развивают листву на разной высоте. Растения, развивающие свою листву в разных ярусах, называют межъярусными (или внеярусными) растениями. Обычно среди ярусов сообщества выделяют главный, который определяет условия существования в фитоценозе. В надземной части фитоценоза главным ярусом обычно бывает один из верхних: или верхний древесный в лесах или наиболее сомкнутый травяной на лугах и в степях и т. д. Однако в некоторых случаях главным ярусом (в который входит эдификатор фитоценоза) оказывается нижний, например ярус сфагновых мхов на торфяном болоте или в заболоченных местах. Некоторые ярусы могут в малоизмененном виде входить в состав нескольких фитоценозов. Так, ярус с господством брусники может встречаться в лесу из европейской ели, в лесу из сибирской ели и даже в лесу из обыкновенной сосны. Эти ярусы называются связующими. Представление об их самостоятельности неверно. Во–первых, при их детальном изучении не обнаруживается полного сходства между связующими ярусами в различных фитоценозах. Во–вторых, их самостоятельность объясняется тем, что под пологом разных древесных пород могут создаться сходные условия. Каждый ярус, входящий в состав фитоценоза, оказывает влияние на другие ярусы и в свою очередь подвергается их влиянию. Поэтому фитоценоз необходимо рассматривать как нечто целое, а ярусы фитоценоза – как его структурные части, которые в некоторых случаях могут быть относительно самостоятельными. Так по экологическим условиям местообитания ельник сфагновый ближе к сосняку сфагновому, чем к ельнику–брусничнику или к ельнику–кисличнику. А сосняк сфагновый ближе к ельнику сфагновому, чем к сосняку с лишайниковым покровом. На основе этого финские исследователи, в частности А. Каяндер (Cajander, 1909), считают, что травяно–кустарничковый и мохово–лишайниковый покровы в лесу служат показателями биологической равноценности местообитаний. Такие ассоциации, в которых одинаковы все ярусы, кроме главного (то есть в лесу – кроме древесного), шведские фитоценологи предложили называть ассоциациями–близнецами, а объединение таких ассоциаций–близнецов Н.Я. Кац (Katz, 1929) предложил называть близнецовым рядом. Так, ельник–кисличник, сосняк–кисличник, лиственничник–кисличник представляют собой ассоциации–близнецы, а кисличники в целом – близнецовый ряд. Подземная ярусность фитоценозов изучена хуже, чем надземная. Распределение корней растений по подземным ярусам определяется изменением с глубиной степени увлажнения почвы, ее богатством питательными веществами и уменьшением степени аэрации почвы с глубиной. Подземные ярусы выделяют на основании глубины проникания корней в почву и размещения активной, то есть способной поглощать воду и питательные вещества, части корневых систем, обычно снабженной корневыми волосками. В лесах нередко можно наблюдать три–шесть подземных ярусов. Например, в широколиственном ясенево–дубовом лесу выделяют ярус залегания корневищ и корней мелко укореняющихся трав, ярус корней более глубоко укореняющихся трав, один или два яруса корней кустарников (более поверхностно и более глубоко укореняющихся), причем эти ярусы могут совпадать и тогда объединяются с подземными ярусами трав, два яруса корней деревьев (верхний из них образуют корни ясеня, нижний – более глубоко идущие корни дуба). Подземные ярусы, также как и надземные влияют друг на друга. Корни, образующие верхний подземный ярус, могут перехватывать дождевую воду у растений, корни которых находятся в более глубоких ярусах. В свою очередь корни, составляющие более глубокие подземные ярусы, перехватывают поднимающуюся капиллярно воду у корней верхних подземных ярусов. Принято считать, что корневые системы растений, входящие в тот или иной подземный ярус, используют влагу и питательные вещества тех почвенных горизонтов, в которых расположен этот ярус. Однако в фитоценозах с недостаточным увлажнением в приповерхностных слоях образуются тонкие активные, так называемые эфемерные корни, очень быстро появляющиеся и так же быстро отмирающие при пересыхании этих слоев (рис. 128).
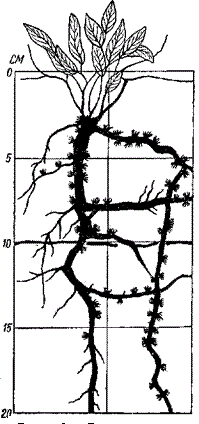
Рисунок 128 – Верхняя часть корневой системы перечника (Lepidiutn) с эфемерными корнями и их основаниями (по Шалыту)
В сообществах с преобладанием засухоустойчивых растений зачастую масса корней во много раз больше массы надземных частей растений. Нередко корневые системы смыкаются в таких сообществах, где надземные части растений разобщены. Иногда, например, на щебнистых склонах, происходит дифференциация корневых систем: образуются корни абсорбирующие и прикрепляющие. Сложен вопрос о ярусном распределении водных растений: одни из них свободно плавают на поверхности воды (нейстон) или в ее толще (планктон), другие прикрепляются к поверхности грунта и не имеют корневой системы как таковой, третьи укореняются в грунте водоема. Так, например, ассимилирующие органы белой кувшинки и малой ряски находятся на поверхности воды. Но слоевища ряски свободно плавают на поверхности, а листья кувшинки отходят от корневищ, находящихся в толще грунта. Вегетативные органы пузырчатки, рдеста блестящего и харовой водоросли могут находиться на одной глубине в толще воды, но пузырчатка – свободноплавающее растение, рдест имеет корневище в толще грунта, а харовая водоросль прикрепляется к поверхности грунта.Поэтому для водных растений следует выделять ярусы прикрепленных видов, указывая глубины, на которых располагаются листья растений, образующих эти ярусы, и ярусы свободноплавающих видов. Некоторые планктонные организмы, не связанные с определенными глубинами, занимают в фитоценозах положение, аналогичное тому, которое занимают межъярусные эпифиты в наземных фитоценозах. В основном это микроскопические водоросли. Животные также преимущественно приурочены к тому или иному ярусу растительности. Некоторые из них вообще не покидают соответствующего яруса. Например, среди насекомых выделяют следующие группы: обитатели почвы – геобий, наземного, поверхностного слоя – герпетобий, мохового яруса – бриобий, травостоя – филлобий, более высоких ярусов – аэробий. Среди птиц есть виды, гнездящиеся только на земле (куриные, тетеревиные, коньки, овсянки и др.), другие – в кустарниковом ярусе (певчие дрозды, снегири, славки) или в кронах деревьев (зяблики, корольки, щеглы, крупные хищники и др.). Таким образом, ярусность представляет собой одну из основных особенностей фитоценоза, возникшую в процессе естественного подбора различных форм для совместной жизни. Горизонтальная структура. Мозаичность. Комплексность. Обычно фитоценоз не бывает совершенно одинаковым на всем своем протяжении. В лесу отличают участки под кронами деревьев и более осветленные, на болотах – кочки и пространства между кочками, в степях – дерновины ковыля со всеми связанными с ними растениями и участки между ними (дерновинами) и т. д. Такие участки, различающиеся друг от друга по особенностям строения растительного покрова, называются микроценозами (Быков, 1953), или микрофитоценозами (Лавренко, 1959). А. А. Гроссгейм (1929) называл их микроассоциациями, П. Д. Ярошенко (1953, 1961) – микрогруппировками, а Н.В. Дылис – парцеллами. Все эти структурные части сообщества, а также вертикальные – ярусы, биогеогоризонты, и более мелкие структурные образования, можно назвать и ценоэлементами. Ценоэлемент – неконкретизированное обозначение любой структурной части или элемента растительности (Трасс, 1970). В биогеоценологии наибольшее признание при изучении горизонтальной неоднородности ценозов получила парцелла как основной ценоэлемент. Этот термин был предложен учеником В.Н. Сукачева Н.В. Дылисом (1969, 1978 и др.). Парцеллы – это структурные части горизонтального расчленения сообщества, обособленные друг от друга на всю вертикальную толщу сообщества (то есть включающие не только растения, но и животных, почвы особенности нано рельефа и вариации микроклимата) (Дылис, Уткин и Успенская, 1964).Парцеллы различаются между собой составом, структурой, свойствами компонентов (лесообразующих пород и растений нижних ярусов), распределением их по площади, количественными показателями. В тоже время между ними (компонентами) существует связь, главная роль в которой принадлежит эдификаторам, то есть тем видам и ярусам, которые обладают самыми сильными средообразующими свойствами. В практике лесоведения и фитоценологии представление о парцелле ассоциируется с составной частью фитоценоза – самого важного и более доступного для изучения звена биогеоценоза. Каждому типу леса присущ свой набор парцелл (рис. 129), чередующихся в определенной последовательности и отражающих как экологические условия место произрастания (влаго – и теплообеспеченность почв, микрорельеф, положение в рельефе, химический состав почвообразующих пород), так и внутриценотическую обстановку (затенение почвы и нижележащих ярусов верхними, конкуренция за питательные вещества и влагу, характер обсеменения, аллелопатия и др.).
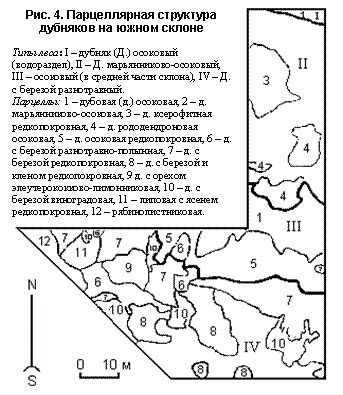
Рисунок 129 – Парцеллярная структура дубняков на южном склоне
При изучении горизонтальной неоднородности напочвенного покрова следует пользоваться термином микрогруппировка. Микрогруппировка – однородная группировка растений напочвенного покрова без разделения его на ярусы, то есть это мелкие сочетания растений внутри сообщества, занимающих одну территорию. Горизонтальная неоднородность фитоценоза характеризуется двумя понятиями – мозаичностью и комплексностью. Мозаичность – горизонтальное расчленение внутри фитоценоза, обусловленное ценотическими факторами. Причины ее разные: 1. Неравномерность условий существования, вызванная жизнедеятельностью тех или иных растений: различия в затенении, химизме и физических особенностях опада, в нанорельефе. 2. Результат деятельности животных, выбрасывающих на поверхность почву из глубоких горизонтов (землерои) или нарушающих в некоторых участках обычный строй растительного сообщества (грызуны). Так, например, деятельность животных может иногда определять особенности ландшафта. Настоящие плотины устраивают бобры (рис. 130). Крупные животные–норники, такие, как суслики или сурки, обеспечивают мозаичность растительного и почвенного покрова, так как за счет выбросов почвы формируется микрорельеф, перераспределяющий осадки и видовой состав растений (рис. 131).

Рисунок 130 – Средообразующая деятельность бобров

Рисунок 131 – Роющая деятельность норных животных в степи. Слева направо: перевязка, безногая ящерица желтопузик, слепыш, обыкновенный хомяк, степная пеструшка, европейский суслик, байбак.
3. Изменения среды под влиянием жизнедеятельности отдельных видов растений создают фитогенную мозаичность. Например, она хорошо выражена в смешанных хвойно–широколиственных лесах. По сравнению с лиственными породами ель сильнее притесняет поверхность почвы, задерживает поверхность почвы, задерживает кронами больше дождевой влаги и снега. 4. Способом роста тех или иных растений, образующих кочки или куртины. Так, высокие кочки, образуемые осокой дернистой на болотах, способствуют возникновению весьма разнообразных микроценозов на вершинках, на откосах и в понижениях между кочками. 5. Нередко мозаичность развивается одновременно с развитием фитоценоза или несколько отстает в развитии от фитоценоза и появляется позже его возникновения. Наконец, она может явиться результатом смыкания ранее разобщенных микроценозов, образующих мозаичный фитоценоз постепенно путем слияния (Ярошенко, 1961). Мозаичность – горизонтальное расчленение внутри фитоценоза – не всегда легко отличить от комплексности – сочетания участков различных сообществ, то есть комплексность отражает влияние экологических (экотопических) факторов.П. Д. Ярошенко предложил различать мозаичность и комплексность по следующим признакам: при мозаичности каждое пятно представляет один единственный микроценоз, а при комплексности каждое пятно в свою очередь может состоять из микроценозов, т. е. комплексность – более сложное явление, чем мозаичность. Присутствие общего эдификатора в сообществе, оказывающего определяющее влияние на жизнедеятельность остальные виды, – наиболее надежный признак, по которому мозаичность отличается от комплексности. Остальные перечисленные признаки не всегда оказываются в наличии. Мозаичность может быть неясной (всегда в одновидовых фитоценозах и на начальных стадиях развития многовидовых фитоценозов), и отчетливой (в многовидовых фитоценозах). Последняя бывает плавной, с нерезкими границами микроценозов, и контурной, с относительно резкими границами. Вероятно, нет ни одной категории явлений, характеризующих структуру фитоценоза, представления о которых были бы столь резноречивыми, как синузии. Понятие « синузия » было введено в науку X. Гамсом (Gams, 1918).Синузия рассматривается как структурная часть фитоценоза, характеризующаяся строго определенным видовым составом, а главное, одинаковой жизненной формой входящих в нее видов, то есть эколого–биологическим единством. Это, например, синузия сосны, синузия брусники, синузия зеленых мхов и другие синузий лесной зоны. Синузия – всегда одноярусное образование. Если она состоит из растений одного вида, то называется простой; если из двух и более – то сложной (рис. 132). Синузия, по Гамсу, есть совокупность (объединение, группа) особей одного вида (синузия первого порядка) или сходных видов (синузии второго и третьего порядков). Синузия второго порядка – совокупность особей разных видов, относящихся к сходной жизненной форме. Виды, образующие синузию второго порядка, должны расти совместно и регулярно встречаться в данном сочетании. Например, кленово–дубовый древостой широколиственного леса. Синузия третьего порядка – совокупность видов, совместно образующих либо ярус, либо одноярусное сообщество. Виды, входящие в ее состав, относятся к различным классам жизненных форм. Относительная самостоятельность таких синузий подтверждается и тем, что данный ярус или сочетание ярусов может выступать и отдельно как фитоценоз. Например, мохово–кустарничковая синузия соснового леса способна существовать на пустоши самостоятельно, хотя облик ее будет несколько иным, чем под пологом леса.

Рисунок 132 – Синузиальная структура напочвенного покрова разнотравно–хвощового лиственичника. Синузии: 1 – вейниково–хвощовая; 2 – вейниково–хвощовая с линнеей; 3 – гераниево–хвощовая; 4 – хвощово–вейниковая; 5 – травяно–зеленомошная; 6 – хвощово–брусничная; 7 – княженичная; 8 – хвощево–зеленомошная; 9 – линнеевая; 10 – маршанциевая; 11 – вейниково–хвощовая с майником.
Популяции отдельных видов (синузии первого порядка) могут составить синузию второго порядка (например, мхов, лишайников и кустарничков) и вместе с другой синузией второго порядка (например, травянистых мезофильных и гемикриптофитов, того же размера, что и виды в синузии второго порядка) – синузию третьего порядка. Виды, образующие синузию третьего порядка, должны регулярно встречаться в данном сочетании при сходной обстановке. Таким образом, синузии могут быть диффузно смешанными в одном и том же пространстве. Неопределенность понятия синузии в дальнейшем вызвала различные его толкования. Т. Липмаа объединял синузии, сходные по признакам: ядро характерных видов, определенные состав жизненных форм и экология, в один унион. Таким образом, унион – это типологическая единица синузии. Синузии и унионы свойственны гомогенной и относительно стабилизированной растительности, т. е. находящейся в равновесии с условиями местообитания. В. Н. Сукачев определял синузии следующим образом (1957): синузии – это «структурные части фитоценозов, характеризующиеся определенным видовым составом, определенным экологическим характером видов, их составляющих, пространственной или временной обособленностью, а следовательно, и особой микросредой, создаваемой растениями данной синузии». Сходное определение дает и А. П. Шенников (1964): «Такие структурные части фитоценоза, отграниченные в пространстве (занимающие особую экологическую нишу), отличающиеся одна от другой в морфологическом, флористическом, экологическом и фитоценотическом отношениях, называют синузиями». Несколько иначе рассматривает их Н. А. Миняев (1963). Он указывает, что синузия есть простейшая растительная группировка, характеризующаяся конкурентными отношениями между особями, входящими в ее состав. Б. Н. Норин (1965) полагает, что «синузией будет пространственно обособленная, однородная, одноярусная, сомкнутая в надземных или подземных частях группировка растений». Таким образом, в понимании синузии намечаются два подхода: синузия – понятие экологическое, оно охватывает растения, относящиеся к одной жизненной форме в сообществе; синузия – понятие структурное, ценотическое. Сторонники последней точки зрения понимают под синузией либо любую хоть сколько–нибудь обособленную часть сообщества (Сукачев и его последователи), либо только те структурные части, которые сложены экологически однородными растениями. Последнюю точку зрения развивают Н. А. Миняев, по–видимому, Б. Н. Норин и особенно последовательно П. Д. Ярошенко (1961). Синузией могут быть: ярус, составленный растениями, относящимися к одной жизненной форме, если он однороден на всей территории, занятой фитоценозом; часть яруса в пределах участка, занятого микрофитоценозом, если весь ярус неоднороден и в различных микрофитоценозах, образующих данное сообщество, представлен растениями, относящимися к различным жизненным формам; совокупность эпифитов, относящихся к одной жизненной форме в пределах всего фитоценоза, если отсутствуют вариации в их экологических особенностях или на протяжении микроценоза, если в разных микроценозах эпифиты представлены разными жизненными формами; совокупность растений, обусловливающих аспект, характеризующихся развитием в одни и те же сроки; в некоторых случаях (в одноярусных фитоценозах) синузия может совпадать с микрофитоценозом. Таким образом, приобретают определенность и понятие «синузия» и понятие «микрофитоценоз». Экологическая структура биоценоза. Каждый биоценоз слагается из определенных экологических групп организмов. Эти группы, занимая сходные экологические ниши, в разных биоценозах могут иметь неодинаковый видовой состав. Например, в сухих аридных условиях доминируют склерофиты и суккуленты, а на увлажненных территориях – гигрофиты.Экологическую структуру биоценоза отражает также соотношение групп организмов, объединяемых сходным типом питания. Так, в лесах преобладают сапрофаги, в степных и полупустынных зонах – фитофаги, в глубинах Мирового океана – хищники и детритоеды и т. д. Наиболее ярко различия в экологической структуре биоценоза проявляются при сравнении сообществ организмов в сходных биотопах разных регионов. Многие виды, определяющие экологическую структуру сообщества, занимают сходные экологические ниши и выполняют одни и те же функции. Они называются замещающими или викарирующими. Например, антилопы в саваннах Африки, бизоны в прериях Америки и кенгуру в саваннах Австралии, куница в европейской и соболь в азиатской тайге. В пределах северотаежной подзоны: ель черная на Аляске, ель обыкновенная на Кольском полуострове и лиственница Каяндера на севере Дальнего Востока.Таким образом, экологическая структура биоценоза – это его состав из экологических групп организмов, выполняющих в сообществе определенные функции.Экологическая структура биоценоза в комплексе с видовой и пространственной, а также с особенностями экологической ниши служит его макроскопической характеристикой. В отличие от микроскопической характеристики, дающей представление о связях каждой популяции и вида в сообществе вплоть до мельчайшей детали их экологии, макроскопическая характеристика дает возможность определить свойства того или иного биоценоза, выяснить его устойчивость во времени и пространстве, а также предвидеть последствия изменений, вызванных влиянием антропогенных факторов. Однако оценка биоразнообразия биоценоза в целом по численности видов будет неправильной, если не учитывать размеры организмов. Ведь в биоценоз входят и бактерии, и микроорганизмы, и травы, и деревья. Поэтому необходимо организмы объединять в группировки, близкие по размерам. Здесь можно подходить и с точки зрения систематики (птицы, насекомые, сложноцветные и т. п.), эколого–морфологической (деревья, травы, мхи и т. п.), либо вообще по размерам (микрофауна, мезофауна и макрофауна почв или илов и т. п.). Многоярусные биоценозы, представленные большим количеством видов растений, животных и микроорганизмов, связанных между собой разнообразными пищевыми и пространственными отношениями, называются сложными . Они наиболее устойчивы к неблагоприятным воздействиям. Исчезновение какого–либо вида существенно не отражается на судьбе таких биоценозов. В них происходит лишь незначительная перестройка организации, при которой популяции одного и даже нескольких видов могут заменяться экологически близкими видами, а стабильность сообщества определяется количественной регуляцией численности одних видов другими. В самых сложных биоценозах тропических лесов никогда не наблюдаются вспышки массового размножения отдельных видов. Для простых биоценозов тундр при определенных условиях характерны «вспышки» численности отдельных доминантов. Обычны сильные подъемы численности леммингов, падения или взлеты численности песцов, питающихся леммингами, оленей, оказывающих существенное влияние на растительный покров. Объясняется это тем, что в упрощенном тундровом биоценозе не хватает видов, которые при необходимости могли бы заменить основной вид и выступить в качестве корма и для хищников, и для растительноядных. Концепция экологической ниши Экологическая ниша – это совокупность всех связей вида со средой обитания, которые обеспечивают существование и воспроизведение особей данного вида в природе.Термин экологическая ниша предложил в 1917 году Дж. Гриннелл для характеристики пространственного распределения внутривидовых экологических группировок. Первоначально понятие экологической ниши было близко к понятию местообитание. Однако в 1927 году. Ч. Элтон определил экологическую нишу как положение вида в сообществе, подчеркнув особую важность трофических связей. Г. Ф. Гаузе расширил это определение: экологическая ниша – это место вида в экосистеме.В 1957–1965 годах Дж. Хатчинсон определил экологическую нишу как часть экологического гиперпространства, в которой возможно существование и воспроизведение вида. Предложил понимать под нишей весь диапазон факторов среды, в котором данный вид в течение длительного времени живёт и размножается.
Допустим, что известны верхние и нижние пределы температуры и влажности, приемлемые для существования какого–либо растения. Если эти сведения представить в системе координат, где температура показана по одной оси, а влажность — по другой, то при пересечении прямых, проходящих через границы интервалов, образуется прямоугольник. Он–то и будет областью, в которой может жить данное растение, то есть его экологической нишей.
Если к этой схеме добавить ещё какое–нибудь условие (например, количество питательных веществ в почве), то осей станет уже три, а ниша на графике примет форму трёхмерной (объёмной) фигуры, похожей на ящик.
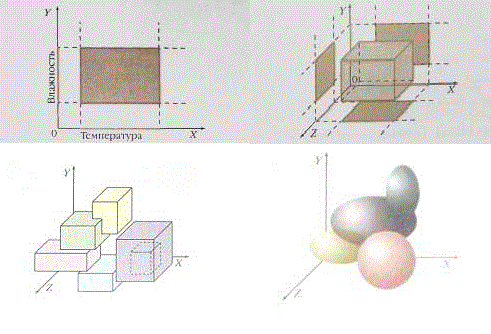
Рисунок 133 – Схема экологической ниши
С помощью такой модели Хатчинсона можно сравнить ниши разных видов организмов. Чтобы мысленно представить эту картину, вообразим, пустую комнату, в которой как бы парят отдельные «ящики» — ниши. Они могут соприкасаться друг с другом, перекрываться или даже вкладываться друг в друга, если условия существования двух видов частично совпадают.
Конечно, такая схема сильно упрощена. Во–первых, ни один организм не выдержит, если сразу несколько факторов имеют предельные значения (именно такие условия создаются в углах «ящика»). Так что в действительности ниши видов имеют сглаженные углы и по форме скорее напоминают сферы. Кроме того, не все сочетания условий для организмов в равной степени хороши. Поэтому реальные ниши «заполнены» неодинаково: где–то численность организмов выше, а где–то — ниже.
Добавив к этой схеме другие показатели, например освещённость или кислотность почвы, можно получить сложные многомерные модели для целых сообществ организмов. Для животных обычно используют три основные оси многомерной ниши. Первая ось показывает пространственную нишу данного вида, т.е. место, где он живёт. Вторая ось характеризует пищевую (трофическую) нишу, то есть то, чем вид питается. Третья ось — это временная ниша, отражающая время суток или сезон, когда вид активен. С помощью трёх основных составляющих можно описать положение отдельных видов в разных сообществах и их взаимоотношения.
В 1984 г. С. Спурр и Б. Барнес выделили три компонента ниши: пространственный (где), временной (когда) и функциональный (как). В этой концепции ниши подчеркивается важность как пространственного, так временного компонента ниши, включающего ее сезонные и суточные изменения с учетом цирканных и циркадных биоритмов. Часто используется образное определение экологической ниши: местообитание – это адрес вида, а экологическая ниша – его профессия (Ю. Одум). В разных сообществах одну и ту же «профессию» могут иметь разные виды. Например, нишу крупных травоядных в Африке занимают антилопы, в Австралии — кенгуру, а в России — лоси.Одни виды всеядны и живут в разных местообитаниях, другие предпочитают только определённую пищу или условия среды. Разнообразие ресурсов, используемых данным видом, называют шириной ниши. Чем шире ниша, тем больше размеры «ящика» в модели Хатчинсона. Например, сумчатый медведь коала питается только молодыми листьями определённых видов эвкалиптов (из–за такой разборчивости коала очень трудно содержать в зоопарках), т.е. он занимает узкую (специализированную) пищевую нишу. А вот у виргинского опоссума пищевая ниша очень широка: этот всеядный зверёк ест практически любую пищу. Многочисленными экологическими исследованиями специализация выгоднее в постоянной среде, а в изменчивых условиях надёжнее иметь широкую нишу. Ресурсов, как правило, на всех не хватает. Поэтому в природе всё время идёт явное или скрытое соперничество за пищу, за место. Самая ожесточённая конкуренция возникает между видами, живущими в одном местообитании, питающимися одинаковой пищей, активными в одно и то же время, то есть, занимающими похожие экологические ниши. В итоге более сильный конкурент вытеснит своего соперника. Этот принцип конкурентного исключения сформулировал и подтвердил экспериментами российский учёный Г. Ф. Гаузе. На опытах с инфузориями и дрожжами Гаузе доказал, что при полном перекрывании экологических ниш один вид довольно быстро вытесняет другой. Поэтому виды, живущие в одном сообществе, всегда занимают несовпадающие ниши. Например, в африканской саванне вместе живут разные копытные животные. Все они травоядные, но их пищевые ниши различаются: жирафы едят листву, антилопы – молодые побеги кустарников и верхушки высоких трав, зебры – траву, а бегемоты – водную растительность. Разные виды ящериц охотятся на разную добычу, в разных местах (на земле, среди камней, на ветках кустов и деревьев) и в разное время суток, т.е. их ниши могут различаться по всем трём основным измерениям. Такой способ «упаковки» ниш называется комплементарным (взаимодополняющим). В 1859 г. английские колонисты завезли в Австралию 24 диких европейских кролика. Спустя несколько десятилетий длинноухие зверьки стали настоящим бичом Зелёного континента: они уничтожали всё, что только могло быть съедено: и естественную растительность, и посевы на полях фермеров. Для борьбы с кроликами завозили лис, применяли яды, испробовали даже бактериологическое оружие. Но и по сей день единственное, чего удалось достичь, – это стабилизировать численность зверьков. В начале XX в. из Северной Америки в Европу с грузом картофеля был случайно завезён золотисто–полосатый колорадский жук. Не прошло и столетия, как, размножившись в невероятных количествах, этот вредитель картофеля завоевал гигантские территории от Атлантики до Урала: никакие агротехнические ухищрения и ядохимикаты не могут остановить его победоносного шествия. Довольно и двух примеров, чтобы понять, какое огромное практическое значение имеют исследования экологии популяций – совокупностей особей одного вида (будь то кролик, колорадский жук, ель или орхидея), занимающих определённую (обычно ограниченную) территорию. Таким образом, чтобы охарактеризовать экологическую нишу вида, необходимо знать не только, где он живет, но и чем он питается, чьей добычей является, каким образом и где отдыхает и размножается, способен ли он к передвижению и, наконец, как он воздействует на другие элементы биогеоценоза. В разных частях света и на разных территориях встречаются неодинаковые в систематическом отношении, но экологически сходные виды. Например, травянистая и лесная растительность Австралии по видовому составу существенно отличается от растительности сходного климатического района Европы или Азии, но растения–продуценты в и там и там выполняют аналогичную функцию. То же можно сказать и о сосне в борах и дубе в дубравах. Это касается также антилоп Африки и кенгуру Австралии. Сумчатый волк (Thylacinus cynocephalus) в Тасмании и обыкновенный волк в Европе тоже ведут одинаковый образ жизни, обладают сходным типом питания. То есть в различных биогеоценозах и продуценты и консументы выполняют одинаковые функции, занимают одинаковые экологические ниши. Вместе с тем часто близкие в систематическом отношении виды, поселяясь рядом в одном биоценозе, занимают разные экологические ниши (рис. 134). Это приводит к уменьшению конкурентного напряжения между ними. У растений, живущих в одном ярусе, экологические ниши сходны, а у растений разных ярусов – различные экологические ниши, что ослабляет конкуренцию между ними. Например, сосна и черника в бору растут в разных ярусах и занимают разные экологические ниши. Такие водные растения, как рдесты (Potamogeton), кубышка (Nuphar luteum), водокрас (Hydrocharius morsus), ряски (Lemnaceae), поселяются вместе, но распределяются по различным нишам.
Рисунок 134 – Экологические ниши организмов, питающихся корнями (1), корневыми выделениями (2), листьями (3), тканями стебля и ствола (4), плодами и семенами (5,6), цветками и пыльцой (7,8), соками (9) и почками (10)
Один и тот же вид в разных условиях может по–разному реагировать на факторы среды, то есть тоже распределяются по разным эконишам. Так, черника и седмичник (Trientalis europaea) в лесах умеренной полосы являются типичными теневыми формами, а в тундре и лесотундре растут на открытых пространствах и становятся световыми Кроме того, в разные периоды развития и даже в разные сезоны года один и тот же вид может занимать различные экологические ниши. Например, головастик питается растительной пищей, а взрослая лягушка – типичное плотоядное животное, поэтому им свойственны различные экологические ниши и специфические трофические уровни. Разными экологическими нишами зимой и летом в связи с миграциями характеризуются и перелетные птицы; у насекомоядных птиц зимние экологические ниши отличаются от летних. Гусеницы бабочек, личинки пилильщиков, мух, жуков и взрослые насекомые также занимают различные экологические ниши. Среди водорослей имеются виды, которые функционируют то, как автотрофы, то, как гетеротрофы. Существует понятие фундаментальной ниши (теоретически возможной ниши для вида при отсутствии конкуренции) и реализованной ниши (фактической, в конкретных условиях). Понятие ниши тесно связано с понятием экологического оптимума. Фундаментальной нише соответствует аутэкологический (или физиологический) оптимум, то есть оптимум в отсутствие конкурентов. Реализованной нише соответствует синэкологический оптимум, то есть оптимум в конкретных условиях, в присутствии всего комплекса биотических и антропогенных факторов. Положение популяции в экосистеме может быть различным: от полного доминирования (сосна обыкновенная в сосновом лесу) до полной зависимости и подчиненности (светолюбивые травы под пологом леса). При этом она, с одной стороны, стремится максимально полно осуществлять свои процессы жизнедеятельности в своих интересах, а с другой – автоматически обеспечивает жизнедеятельность других популяций этого же биоценоза, являясь компонентом цепи питания, а также через топические, приспособительные и иные связи. Таким образом, каждая популяция, как полноправный представитель вида в экосистеме, имеет в ней свое место. Американский эколог Р.Макинтош назвал его экологической нишей. Основные компоненты экологических ниш: 1.Определенное местообитание (физико–химические свойства экотопа и климатические условия); 2. Биоценотическая роль (производитель, потребитель или разрушитель органического вещества); 3. Положение внутри собственного трофического уровня (доминирование, содоминирование, подчиненность и т.п.); 4. Место в пищевой цепи; 5. Положение в системе биотических отношений. Динамика экологических ниш. В естественных условиях никакая популяция или особь не находятся сколь–либо длительное время в идеальных условиях. Реальные условия существования определяют параметры действительной сферы жизнедеятельности и формируют так называемую реализованную экологическую нишу. Конечно, любое сообщество организмов, в том числе отдельные особи, стремятся максимально реализовать свой биотический потенциал и занять лучшее место в собственном трофическом уровне. Представление о наиболее благоприятных условиях жизнедеятельности для них рассматривается как фундаментальная экологическая ниша. Условия внешней среды постоянно, более или менее системно, изменяются, создавая этим лучшие или худшие возможности для жизнедеятельности популяции. Они же (популяции), в свою очередь, расширяют свою экологическую нишу или вынужденно сужают, создавая определенный динамический эффект. Как правило, внутри одного трофического уровня находятся несколько однотипных популяций. Например, продуценты луговых экосистем представлены многими видами трав: тимофеевка луговая (Рleum pretense L.), ежа сборная (Dactylis glomerata L.), клевер луговой (Trifolium pretense L.), одуванчик лекарственный (Taraxacum officinale L.) и др. Однако, даже самый обыкновенный ценоз по годам может значительно изменять свой ботанический состав. В иные годы увеличивается доля одного вида (расширяется его экологическая ниша) в иные – другого вида. Часто наблюдаются случаи, когда внешние условия резко меняются, в пользу одного из видов, который тут же расширяет свою экологическую нишу и становится доминирующим. В большинстве случаев сосуществующие виды в одном трофическом уровне из–за конкурентной неспособности захватить экологические ресурсы, которых часто не хватает, должны смириться с «соседями» и «подвинуться», то есть сместить свою экологическую нишу. В противном случае один из видов будет вытеснен из ценоза. Подобное явление описывается принципом конкурентного исключения Гаузе (1994): два вида не могут устойчиво существовать в ограниченном пространстве экосистемы и смещают свои экологические ниши, если жизнедеятельность их ограничена каким–либо важным ресурсом, количество и (или) доступность которого лимитирована (по Мамедову Н.М. и др., 1996). Например, разные виды цапель (Ardea L.) в одной и той же экосистеме, на одних и тех же прибрежных участках имеют разные приспособленности к поиску пищи: цапля красная взбаламучивает воду, пугая рыбу, затем широко раскрывает крылья, создает тень. Рыбы принимают тень за убежище, устремляются к нему и становятся добычей птице. Другой вид цапель – зеленая кваква – пассивно поджидает проплывающую мимо рыбу. Экологическая ниша какого–либо вида по одному фактору окружающей среды может быть изображена графически следующим образом (рис. 135). Зона пересечения данных прямых может рассматриваться как экологическая ниша по двум факторам только при отсутствии между ними взаимодействия (рис. 136). Если же оно налицо, то экологическую нишу представляют в следующем виде (рис. 137). Чем ближе к ее центру, тем благоприятнее условия существования для данного вида.
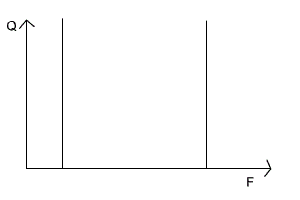
Рисунок 135 Экологическая ниша 1 –
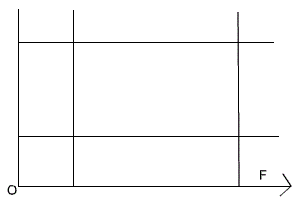
Рисунок 136 Экологическая ниша
2 ( по двум факторам среды) –
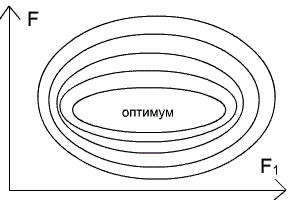 > >
Рисунок 137 Экологическая ниша
3 ( по трем факторам среды) –
Поскольку живые организмы существуют в пределах множества экологических факторов, то реальная экологическая ниша может быть представлена в виде некоего гиперобъёма с зоной оптимума в центре. Гиперобъём фундаментальной экологической ниши будет определяться толерантностью вида по всем факторам, а гиперобъём реализованной экологической ниши – динамически изменяться в пределах фундаментальной. Факторы внешней среды определяют динамику экологических ниш сообществ организмов и отдельных особей как в целом, так и на каждом этапе онтогенеза. Следует учитывать, что на разных этапах онтогенеза особи могут занимать разные экологические ниши. Например, личинки (гусеницы) капустной белянки (Pieridae L.) и взрослое насекомое (имаго) имеют разные способы питания и разные источники пищи, а головастик лягушки и взрослая особь – не только разные источники питания, но и различные среды обитания, относясь к иным трофическим уровням. Итак, изменение условий существования (смена времён года, смена сезонов, антропическое воздействие и т.п.), переход из одного возрастного состояния в другое приводят к динамическим изменениям реализованных экологических ниш, вплоть до их смены. Видовой состав естественных биоценозов близок к полночленному. Однако, в них могут проникать новые виды как в результате акклиматизации человеком, так и не преднамеренно. Попытки изменения видового состава экосистем могут быть удачными при исключительно тщательном анализе всех взаимоотношений в сообществе, а не только при определении свободной экологической ниши по пищевому фактору. Важно учитывать комплекс факторов, в том числе непрерывность и законченность пищевых цепей. Представление об экологических нишах настолько фундаментально для всей природоохранной и природопользовательской деятельности, что Э.Пианка (1971) даже высказал предположение, что всю экологию можно рассматривать как науку об экологических нишах. Суть принципа вытеснения, также известного как принцип Гаузе, состоит в том, что каждый вид имеет свою собственную экологическую нишу. Никакие два разных вида не могут занять одну и ту же экологическую нишу. Сформулированный таким образом принцип Гаузе подвергался критике. Например, одним из известных противоречий этому принципу является парадокс планктона. Все виды живых организмов, относящихся к планктону, живут на очень ограниченном пространстве и потребляют ресурсы одного рода (главным образом солнечную энергию и морские минеральные соединения). Современный подход к проблеме разделения экологической ниши несколькими видами указывает, что в некоторых случаях два вида могут разделять одну экологическую нишу, а в некоторых такое совмещение приводит один из видов к вымиранию.Экологическая ниша не может быть пустой. Если ниша пустеет в результате вымирания какого–то вида, то она тут же заполняется другим видом. Человек как биологический вид занимает свою собственную экологическую нишу. Человек может обитать в тропиках и субтропиках, на высотах до 3–3,5км над уровнем моря. Реально в настоящее время человек живёт в значительно больших пространствах. Человек расширил свободную экологическую нишу благодаря использованию различных приспособлений: жилища, одежды, огня и пр. У каждого вида животных своя ниша, что сводит к минимуму конкуренцию с другими видами. Поэтому в сбалансированной экосистеме присутствие одного вида обычно не угрожает другому. Адаптация к разным нишам связана с действием закона лимитирующего фактора. Пытаясь использовать ресурсы за пределами своей ниши животное сталкивается со стрессом, то есть с ростом сопротивления среды. Иными словами, в собственной нише его конкурентоспособность велика, а вне ее значительно ослабевает или пропадает вовсе. Адаптация животных к определённым нишам заняла миллионы лет и протекала в каждой экосистеме по–своему. Ввезённые из других экосистем виды могут вызвать вымирание местных именно в результате успешной конкуренции за их ниши (рис. 138). Например: Скворцы, завезённые в Северную Америку из Европы, за счёт своего агрессивного территориального поведения вытеснили местных «синих» птиц. Одичавшие ослы потравили пустынные экосистемы, вытеснив оттуда снежного барана. В 1859 году в Австралию из Англии завезли кроликов для спортивной охоты. Природные условия оказались для них благоприятными, а местные хищники – не опасными. В результате кролики расплодились на столько, что уничтожили обширные территории пастбищ. Лисы, привезённые для уничтожения кроликов нашли более лёгкую добычу (местных сумчатых). Лишь позже удалось определить паразита и решить проблему. Земледельцы ищут методы борьбы с сорняком ранее не встречавшимся в нильской долине. Невысокое растение с крупными листьями и мощным корнем уже несколько лет ведёт наступление на обрабатываемые земли Египта. Местные агрономы считают его чрезвычайно активным вредителем. Оказывается, что это растение известно в Европе под названием «хрен деревенский». Вероятно, его завезли русские специалисты, строившие металлургический комбинат. Концепция экологической ниши применима и к растениям. Как и у животных их конкурентоспособность высока лишь в определённых условиях. Например: платаны растут по берегам рек и в поймах, дубы на склонах. Платан приспособлен к переувлажнённой почве. Семена платана распространяются вверх по склону и этот вид может расти там при отсутствии дубов. Аналогично, жёлуди, попадая в пойму, гибнут из–за избытка влаги и не способны конкурировать с платанами. Экологическая ниша человека – состав воздуха, воды, пищи, климатические условия, уровень электромагнитного, ультрафиолетового, радиоактивного излучения и пр.

Рисунок 138 Животные, занимающие экологическую нишу дятла в тех местах, где дятлы отсутствуют (по книге Э. Майра и др. «Эволюция»)
1. Один из галапагосских вьюрков, добывающий насекомых из–под древесной коры при помощи кактусовой колючки. 2. Одна из гавайских цветочниц, у которой в процессе эволюции развился клюв, сходный с клювом дятла. 3. Полосатый поссум с Новой Гвинеи. 4. Руконожка ай–ай – примат с Мадагаскара. Оба этих млекопитающих благодаря развитию удлинённых пальцев на передних конечностях могут извлекать из деревьев личинок насекомых.
|