Тема 4.2. Потоки вещества и энергии в экосистемах
Структура экосистемы характеризуется (как и биогеоценоза) наличием двух системообразующих частей – биотопа и биоценоза, которые включают в себя четыре функционально различных компонента, осуществляющих круговорот веществ и поток энергии. Первый компонент – абиотические факторы среды, или биотоп (местообитание), то есть весь комплекс неживой природы, в том числе: физические факторы (солнечная энергия, давление, теплота и прочее), химические факторы (вода, биогенные элементы, минеральные соли, газы), откуда биоценоз черпает средства жизни и куда выделяет продукты своего обмена. Второй компонент – продуценты, или образователи, представленные комплексом разнообразных зеленых растений и фотосинтезирующих бактерий. Они обеспечивают все живущее здесь население органическим веществом и запасенной энергией. Основная масса из них – автотрофы, то есть фотосинтезирующие макро– и микроорганизмы, частично их дополняют и хемосинтезирующие бактерии. Третий компонент – комплекс всевозможных организмов – гетеротрофов, живущих за счет готовых органических веществ, которые создали продуценты. Это – консументы, или потребители. Среди них – многообразие организмов растительноядных, плотоядных, паразитических и пр. Четвертый компонент – комплекс организмов, разлагающих органические соединения до минерального состояния. Это – редуценты, или разлагатели, представление макро– и микроорганизмами, среди них бактерии, грибы, животные (простейшие). Между четырьмя звеньями существует тесная взаимосвязь и зависимость (рис. 141). Свойства каждого звена во многом определяют свойства каждого последующего звена, равно как и оно само находится в зависимости от функционирования других звеньев. Все процессы жизнедеятельности организмов в биогеоценозе протекают на фоне взаимодействия с окружающей средой. Сюда включается аккумуляция солнечной энергии, потребление химических веществ из почвы, воздуха и возвращение веществ от живых организмов в абиотическую среду. 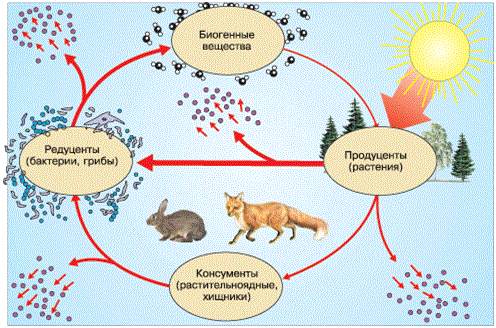
Рисунок 141 – Необходимые компоненты экосистемы
Экосистемы могут быть любого ранга в зависимости от того, какого ранга (то есть какое количество и качество поступающей энергии) в нем осуществляется круговорот веществ, вовлекающий с помощью живого населения (продуценты, консументы и редуценты) биогенные элементы абиотической среды (углерод, азот, фосфор, калий и т. д.) в циклы. Каждая экосистема характеризуется своим особым биогеохимическим циклом движения веществ и энергии. В действительности, хотя в каждой экосистеме биогеохимический круговорот (то же на уровне биогеоценоза) может быть завершен, в реальности обособленных круговоротов в природе нет – все они объединяются (на уровне биосферы) в единую систему – биологический круговорот. В этом процессе проявляется глобальная функция живого населения нашей планеты. Таким образом, основными компонентами экосистемы являются: природный резервуар элементов минерального питания и сообщества живых организмов с функционально различными свойствами в виде продуцентов, консументов и редуцентов. Продуценты – это система автотрофов, способных с помощью световой энергии строить ткань своего тела из неорганических соединений. Консументы и редуценты – гетеротрофные организмы. Первые растительноядные и плотоядные организмы, вторые – организмы, разлагающие всевозможные остатки органики. Те и другие в сети цепей питания разрушают органические вещества, созданные продуцентами, возвращают химические элементы в минеральный резервуар неживой природы. В результате вещества вновь становятся доступными для автотрофов. Возврат биогенных элементов в среду осуществляется как в течение жизни организмов (путем дыхания, выделения, дефекации, регулярного отпада частей тела – волос, кожи, рогов, скорлупы, листьев, побегов, корневых волосков и так далее), так и после их смерти в результате разложения трупов и отмерших растений. Известно что ресурсы биогенных элементов, необходимых для построения тел живых организмов на земной поверхности, не безграничны, но система биогеохимических круговоротов придает этим запасам свойство бесконечности, необходимое для продолжения жизни внутри каждой отдельной экосистемы и в глобальном масштабе – в биосфере в целом. Однако следует заметить: в то время как вещество, в том числе биогенные элементы (углерод, фосфор, азот и др.), необходимые для жизни, и вода, многократно используется внутри самой экосистемы, энергия, поступающая в нее извне в виде солнечного излучения, необратимо превращается в тепло. Энергия в экосистеме может накапливаться в виде химических связей органических веществ, снова высвобождаться или экспортироваться по цепям питания, но она не используется вторично. Важно отметить, что звенья, образующие пищевую (трофическую) цепь, неравнозначны с точки зрения занимаемого места в структуре экосистемы. Поэтому экологи вычленяют не просто звенья в пищевой цепи, а рассматривают их как особые трофические уровни. По их количеству определяют трофико– энергетическую структуру экосистемы. На первом трофическом уровне, естественно, находятся продуценты – зеленые растения, на втором – растительноядные организмы, за ними следуют представители более высокого, третьего уровня – плотоядные организмы, на их основе возникает четвертый уровень – плотоядные второго порядка. В экосистеме, богатой видами, может развиваться пять и даже шесть трофических уровней.
Жизнь на Земле существует за счет солнечной энергии – единственный на земле пищевой ресурс, энергия которого в соединении с углекислым газом и водой рождает процесс фотосинтеза. Солнечная энергия через растения как бы передается всем организмам. Энергия передается от организма к организму, создающих пищевую или трофическую цепь, то есть перенос веществ и заключенной в них энергии от автотрофов к гетеротрофам, происходящий в результате поедания одними организмами других. Число звеньев в ней может быть различным, но обычно их бывает от 3 до 5. Совокупность организмов, объединенных одним типом питания и занимающих определенное положение в пищевой цепи называется трофический уровень. Первый трофический уровень – это продуценты (гелиотрофы, хемотрофы), второй – это растительноядные консументы 1 порядка (фитофаги), третий – плотоядные консументы 2 порядка, питающиеся растительноядными консументами 1 порядка (зоофаги), четвертый – редуценты или деструкторы, разлагают органические вещества до неорганических (рис. 142). Большая часть энергии при переходе с одного трофического уровня на другой, более высокий теряется. Приблизительно потери составляют 90 %: на каждый следующий уровень передается не более 10 % энергии от предыдущего уровня. Так, если калорийность продуцента 1000 Дж, то при попадании в тело фитофага остается 100 Дж, в теле хищника 10 %. Таким образом, входя в экосистему, поток лучистой энергии разбивается на две части, распространяясь по двум видам трофических сетей, но источник энергии общий – солнечный цвет. 
Рисунок 142 – Разрушители мертвой древесины (жук бронзовка и его личинка; жук– олень и его личинка; большой дубовой усач и его личинка; бабушка древоточец пахучий и его гусеница; жук красный плоскотел; многоножка кивсяк; черный муравей; мокрица; дождевой червь)
Различают два вида трофических цепей: цепи выедания или пастбищные, которые начинаются с поедания фотосинтезирующих растений (растения – гусеницы – насекомоядные птицы – хищные птицы), и детритные цепи разложения, которые начинаются с остатков отмерших растений, трупов, экскрементов животных (коровий помет – личинки мух – скворцы – ястребы – перепелятники) (рис. 143). В разных типах экосистем мощность потоков энергии через цепи выедания и разложения различна: в водных сообществах большая часть энергии, фиксированной одноклеточными водорослями, поступает к питающимся фитопланктоном животным и далее – к хищникам и значительно меньшая включается в цепи разложения. В большинстве экосистем суши противоположное соотношение: в лесах, например, более 90 % ежегодного прироста растительной массы поступает через опад в детритные цепи. 
Рисунок 143 – ; Примеры цепей питания: А – цепи выедания; Б – цепи разложения
Принцип биологического накопления. В круговорот веществ в экосистеме часто добавляются вещества, попадающие сюда извне. Эти вещества концентрируются в трофических цепях и накапливаются в них, то есть происходит биологическое накопление. Это явление наглядно видно на примере концентрирования радионуклидов и пестицидов в трофических цепях. Наиболее известна способность к биологическому накоплению ДДТ – вещества, ранее широко применяемого для борьбы с вредными насекомыми и запрещенного к применению в настоящее время. Живые организмы, входящие в состав экосистемы, неодинаково ассимилируют вещества и энергию. Зеленые растения непрерывно усваивают из атмосферного воздуха огромное количество углекислого газа и образуют ежегодно около 170 млрд. тонн органического вещества. При этом растения запасают много солнечной энергии, общее количество которой оценивается в 20,9*1022 кДж. Кроме того, синтез органического вещества может осуществляться и бактериями. Животные в отличие от растений и бактерий не могут осуществлять реакции фото– и хемосинтеза и вынуждены использовать солнечную энергию опосредованно – через органическое вещество, созданное фотосинтезирующими организмами. Таким образом, благодаря разнообразию населения в биогеоценозе образуется цепь последовательной передачи вещества и заключенной в нем энергии от одних организмов к другим. Этот сквозной поток энергии, проходя через трофическую цепь экосистемы, постепенно гасится, поскольку на всех этапах этой цепи идет потеря энергии, главным образом в виде тепла. Количественное выражение вещественно– энергетических потоков может служить основой для оценки эффективности продукционных процессов и средообразующих функций ценоза. Для этой цели разработано множество показателей. Ниже дается трактовка основных терминов, используемые при рассмотрении изменений биомассы и биологической продукции. Различают общую, первичную и вторичную продукцию. 1. Общая продукция (биомасса) – запасы всей органической массы, т.е. масса организмов, присутствующих в экосистеме в момент наблюдения и учтенная на единицу площади. Легче всего поддается учету и измерению запасы биомассы и продукция живых растений – автотрофов, она называется фитомассой. Зоомассой, соответственно, называется масса консументов. Все "массы" могут быть выражены в единицах массы и энергии (калориях). 2. Первичной продукцией, или брутто– продукцией, называется органическая масса, создаваемая продуцентами в единицу времени. Первичная валовая продукция (Пв) – суммарная продукция фотосинтеза (суммарная ассимиляция), включающая, следовательно, и вещество, сжигаемое при дыхании за единицу времени. Жизнь на Земле существует за счет солнечной энергии – единственный на земле пищевой ресурс, энергия которого в соединении с углекислым газом и водой рождает процесс фотосинтеза. Фотосинтез происходит только в растениях (рис.144). В ходе фотосинтеза растение вырабатывает из неорганических веществ необходимые для всего живого органические вещества. Диоксид углерода, содержащийся в воздухе, проникает в лист через особые отверстия в эпидермисе листа, которые называют устьицами; вода и минеральные вещества поступают из почвы в корни и отсюда транспортируются к листьям по проводящей системе растения. Энергию, необходимую для синтеза органических веществ из неорганических, поставляет Солнце; эта энергия поглощается пигментами растений, главным образом хлорофиллом. В клетке синтез органических веществ протекает в хлоропластах, которые содержат хлорофилл. Свободный кислород, также образующийся в процессе фотосинтеза, выделяется в атмосферу. 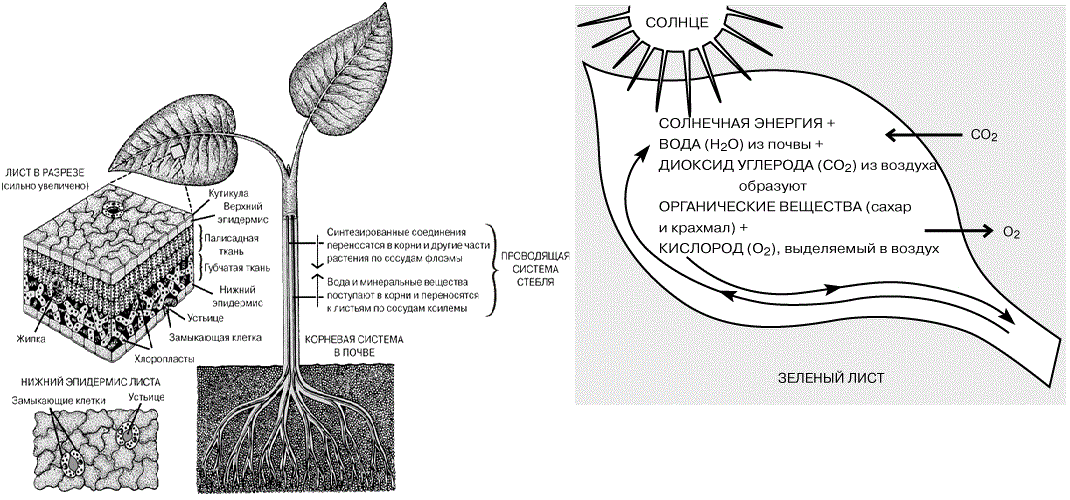
Рисунок 144 – Схема фотосинтеза
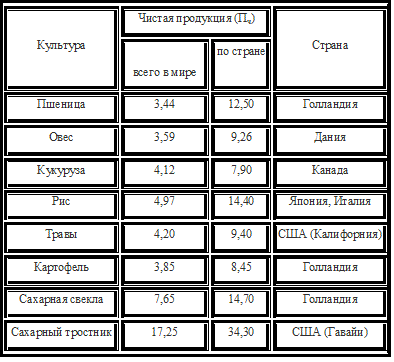
Даже при оптимальных условиях растения на суше могут использовать лишь несколько процентов видимого излучения Солнца, а коэффициент полезного действия для всей поверхности суши составляет 0,1 – 0,3 %. Чистая первичная продукция (Пч), или нетто– продукция, – это суммарная продукция фотосинтеза за вычетом вещества, затраченного растениями на дыхание, т.е. вещество, которое можно взвесить при уборке урожая (табл.). Определяется эффективностью фотосинтеза, площади фотосинтезирующих органов, а также от длительности периода активного фотосинтеза. Для многолетних растений Пч – масса годичного прироста. Таблица 12 Значения чистой первичной (годовой) продукции некоторых культур (растения целиком), т/га абс.– сух вещества
Согласно Ю. Одуму (1959), значения первичной продукции для разных типов биогеоценозов следующие: 1. Очень низкой продуктивностью обладают океанические глубины и пустыни (0,1 г/м2 в день). Хотя ограничивающие факторы двух сред совершенно различны (в океанах – питательные вещества, в пустынях – вода), как различны и продуцирующие организмы (водоросли – в океанах), глубины океанов и аридные территории по продуктивности можно рассматривать как пустыни. 2. Сходной низкой продуктивностью (0,5 – 3 г/м2 в день) характеризуются травяные формации, морские литорали и плохо культивированные земли. 3. Продуктивностью от 10 до 20 г/м2 в день обладают влажные леса, земли с интенсивным сельскохозяйственным использованием, аллювиальные формации, эстуарии и коралловые рифы. Максимально возможная продуктивность экосистемы, по– видимому, не может превышать 25 г/м2 в день. Следовательно, разные биогеоценозы продуцируют неодинаковое количество органических веществ. Мало их образуется в пустынных экосистемах и тундрах. Наиболее продуктивны леса, особенно тропические (рис. 145). 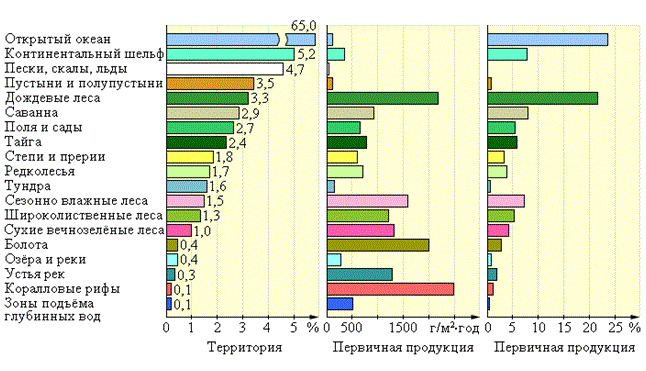
Рисунок 145 – Первичная продукция в различных природных зонах Земли
3. Вторичная продукция (П2) – прирост за единицу времени массы консументов. 4. Опад – вся отмершая масса растений, лежащая на поверхности – мелкие растительные остатки, характеризующиеся поэтому более быстрыми в обычных условиях темпами разложения (листву, генеративные органы многих растений, ветви и побеги толщиной менее 5 мм, травы, мхи и лишайники, кусочки коры и др.) 5. Отпад – отмершие органы растений – более крупные, чем опад. Обычно сильно лигнизированные остатки (стволы, ветви толщиной более 5 мм, шишки хвойных пород, крупные куски коры деревьев). 6. Величина годичного опада и отпада органического вещества растительного происхождения, определяемая по массе и энергии и с учетом ярусного сложения фитоценозов. 7. Годичная норма разложения растительных остатков как результат деятельности сапрофагов и редуцентов и физико– химических процессов в слое лесной подстилки. 8. Фракционный состав запаса и годичного прироста фитомассы; выражается процентным соотношением массы разных фракций фитомассы: стволы, ветви и побеги, листва, генеративные органы, корни. В пределах этих групп фракций желательна дальнейшая дифференциация: для стволов, ветвей и корней – на древесину и кору; для ветвей и корней – по группам толщины; для хвои вечнозеленых пород – по возрастам; для генеративных органов с целью полного учета общего годичного прироста – на цветки (или стробилы); недоразвившиеся плоды (или женские стробилы) – на семена и другие части плодов (или женских стробил) в зрелом состоянии 9. Соотношение массы надземных частей растений к массе корней, определяемое: а) в запасе; б) в годичном приросте фитомассы. 10. Отношение суммарной длины древесных частей растений в надземном слое к суммарной длине корней, если последний показатель будет определен. 11. Коэффициенты связи между весом разных частей деревьев и таксационными признаками стволов. Эти показатели, в зависимости от принятого метода обработки данных модельных деревьев, могут быть представлены либо значениями постоянных коэффициентов аллометрических уравнений (регрессий), либо средними значениями относительных коэффициентов, получаемых при делении веса разных частей деревьев на d2 или на d2h (Ватковский, 1968), где d – диаметр ствола на высоте груди (в см), h – высота ствола (в м). 12. Коэффициент продуктивности (или эффективности работы) ассимиляционного аппарата растений, выражается отношением общего годичного прироста (нетто– продукции) к массе листвы. 13. Индекс листовой поверхности LAI (общий), или отношение площади поверхности листвы (для листьев берется односторонняя поверхность, для хвои – общая) отдельных ярусов и фитоценозов в целом к площади участка в 1 га.. 14. Коэффициент эффективности работы трофического уровня продуцентов; определяется по отношению энергетических эквивалентов для общего и чистого годичных приростов фитомассы к количеству солнечной энергии, поступающей в систему в течение года и (отдельные расчеты) за вегетационный период. 15. Отношение массы листвы к массе всасывающих корней (в расчете на 1 га). Перечисленные и многие другие продукционные показатели отражают разные стороны жизнедеятельности биогеоценоза, его географические, экотопические и возрастные особенности, взаимоотношения между компонентами в борьбе за жизненные факторы (влагу, тепло, свет). Во многом они сходны, в первую очередь сходны тенденции изменения разных показателей, в то же время могут существенно различаться. Так, в зависимости от указанных выше факторов, в лесных биогеоценозах сильно варьирует структура древостоя. Доля стволовой массы может колебаться от 40– 50 до 80– 85 %. В условиях тайги приближенная доля средних значений участия стволовой древесины в фитомассе древостоя колеблется примерно от 45 до 60 %; доля крон – от 12 до 35 %; доля корней – от 11 до 22 %. Энергетическая эффективность животных Путь каждой конкретной порции энергии, накопленной зелеными растениями, короток. Она может передаваться не более чем через 4– 6 звеньев ряда, состоящего из последовательно питающихся друг другом организмов (рис. 146). 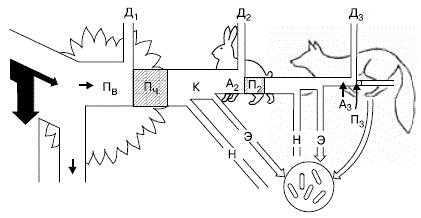
Рисунок 146 – Поток энергии через три уровня простой пищевой цепи (по П. Дювиньо и М. Тангу, 1968): Пв – продукция валовая; Пч – продукция чистая; К – продукция, использованная на корм; А2, А3 – корм, ассимилированный консументами; н – неиспользованная часть продукции; П2 – вторичная продукция (травоядные); П3 – прирост хищников; Д1– Д3 – траты энергии на обмен веществ (траты на дыхание) на разных уровнях пищевой цепи
Энергетический баланс консументов складывается следующим образом. Поглощенная пища обычно усваивается не полностью. Неусвоенная часть вновь возвращается во внешнюю среду (в виде экскрементов) и в дальнейшем может быть вовлечена в другие цепи питания. Процент усвояемости зависит от состава пищи и набора пищеварительных ферментов организма. Пища животного происхождения переваривается легче, чем растительная пища. Эффективность ассимиляции у хищных видов составляет от 60 до 90% потребленной пищи, причем виды, питающиеся насекомыми, стоят на нижнем конце этого ряда, а виды, питающиеся мясом и рыбой, – на верхнем. Жесткий хитиновый наружный скелет, составляющий значительную часть веса у многих видов насекомых, не переваривается, что снижает эффективность ассимиляции у животных, которые питаются насекомыми. Большое влияние на питательную ценность растительной пищи оказывает относительное содержание в ней целлюлозы, лигнина и других неперевариваемых материалов. Стволы и ветви деревьев состоят главным образом из целлюлозы и лигнина, в которых нет азота и многих необходимых минеральных веществ. Жуки– пилилщики, питающиеся деревянистыми частями растений, либо очень медленно растут, либо поедают неодревесневшие живые клетки, лежащие непосредственно под корой. Листья содержат 2– 4% белка и, таким образом, служат более подходящей пищей для растительноядных животных, хотя растения выработали много механизмов для защиты своих листьев, в том числе и разнообразные токсины. В листьях дуба и ряда других деревьев содержатся танины, препятствующие перевариванию заключенных в этих листьях белков растительноядными животными. Семена – наиболее привлекательная растительная пища, потому что они содержат все вещества, необходимые для начала роста растения, и в тоже время представляют собой полноценный корм для поддержания существования растительноядных животных. Сосновые орешки, например, содержат около 50% масел, 30% белка и 5% сахаров. Эффективность ассимиляции у растительноядных животных соответствует питательным свойствам их пищи, достигая 80% при поедании семян, 60% при поедании молодой листвы, 30– 40% при поедании более старых листьев и 10– 20% и даже менее при поедании древесины. Ассимилированная организмом пища вместе с запасом в ней энергии расходуется двояким образом. Большая часть энергии используется на поддержание рабочих процессов в клетках, а продукты расщепления подлежат удалению из организма в составе экскретов (мочи, пота, выделений различных желез) и углекислого газа, образующегося при дыхании. Энергетические затраты на поддержание всех метаболических процессов условно называют тратой на дыхание, так как общие их масштабы можно оценить, учитывая выделение СО2 организмом. Меньшая часть усвоенной пищи трансформируется в ткани самого организма, т. е. идет на рост или откладывание запасных питательных веществ, увеличение массы тела. Эти отношения сокращенно можно выразить формулой: Р = П + Д + Н, где Р – рацион консумента, т. е. количество пищи, съедаемой им за определенный период времени; П – продукция, т. е. траты на рост; Д – траты на дыхание, т. е. поддержание обмена веществ за тот же период; Н – энергия неусвоенной пищи, выделенной в виде экскрементов. Передача энергии в химических реакциях в организме происходит, согласно второму закону термодинамики, с потерей части ее в виде тепла. Особенно велики эти потери при работе мышечных клеток животных, КПД которых очень низок. В конечном счете вся энергия, использованная на метаболизм, переходит в тепловую и рассеивается в окружающем пространстве. Траты на дыхание во много раз больше энергетических затрат на увеличение массы самого организма. Конкретные соотношения зависят от стадии развития и физиологического состояния особей. У молодых траты на рост могут достигать значительных величин, тогда как взрослые используют энергию пищи почти исключительно на поддержание обмена веществ и созревание половых продуктов. Интенсивность питания снижается с возрастом. Так, ежесуточный рацион карпов массой от 5 до 15 г составляет почти 1/4 от массы их тела, у более крупных особей – от 150 до 450 г – всего 1/10, а у рыб массой 500– 800 г – 1/16. Коэффициент использования потребленной пищи на рост (К) рассчитывают как отношение продукции к рациону: К=П/Р, где П – траты на рост, Р – количество пищи, съеденной за тот же период. Двупарноногие многоножки кивсяки в период роста, который продолжается до трех лет, тратят на рост от 6 до 25 % съеденной пищи при усвояемости в среднем 30 %. В последующем их масса стабилизируется. Кивсяки живут до 12 лет. В умеренном поясе они активны 4– 5 месяцев в году. Особь, масса которой во взрослом состоянии 0,5 г, за свою жизнь потребляет 250– 300 г опада (80– 90 г абсолютно сухой массы). Так как кивсяки многократно линяют, часть усвоенной энергии идет на восстановление покровов. Таким образом, отношение съеденного в течение жизни корма к массе взрослого животного составляет 500– 600: 1. Эффективность чистой продукции у животных находится в обратной зависимости от их активности. Деятельность, направленная на поддержание существования, и продукция тепла у теплокровных животных требует расхода энергии, которая могла бы быть использована на рост и размножение. У наземных теплокровных животных в состоянии активности эффективность чистой продукции низкая: у птиц менее 1%, потому что они постоянно находятся в состоянии высокой активности; у мелких млекопитающих с высокой скоростью размножения (например, у кроликов и мышей) – до 6%. Человек поддерживает эффективность чистой продукции крупного рогатого скота на столь высоком уровне, как 11%, забивая животных вскоре после завершения роста или даже раньше. Менее подвижные холоднокровные животные, особенно животные, живущие в воде, затрачивают до 75% ассимилированной ими энергии на рост и размножение, что приближается к максимальной биохимической эффективности роста. У такого гетеротермного животного, как малый суслик, который активен всего 2– 2,5 месяца в году, это соотношение всего около 150: 1. Средний рацион суслика 30 г сухой массы растений (или в среднем 100 г сырой) при массе зверька 200 г и продолжительности жизни 4 года. Постоянно активным в течение года рыжим полевкам нужно гораздо больше энергии для поддержания жизнедеятельности. Взрослые зверьки массой 20 г съедают в среднем до 4 г сухого корма в день. При продолжительности жизни примерно в 24 месяца затрата кормов на жизнь одной особи составляет примерно 30 кг в сырой массе, что приблизительно в 1500 раз больше массы взрослого животного. Эффективность продукции биомассы в пределах одного трофического уровня (эффективность общей продукции) представляет собой произведение эффективности ассимиляции и эффективности чистого роста. Эффективность общей продукции только у нескольких видов теплокровных животный превышает 5%, у птиц не достигает 1%. Эффективность общей продукции у насекомых лежит в пределах от 5 до 15%, а у некоторых водных животных превышает 30%. Эффективность чистой продукции находится в обратной зависимости от эффективности ассимиляции, особенно у водных животных. Таким образом, основная часть потребляемой с пищей энергии идет у животных на поддержание их жизнедеятельности и лишь сравнительно небольшая – на построение тела, рост и размножение. Иными словами, большая часть энергии при переходе из одного звена пищевой цепи в другое теряется, так как к следующему потребителю может поступить лишь та энергия, которая заключается в массе поедаемого организма. По грубым подсчетам, эти потери составляют около 90 % при каждом акте передачи энергии через трофическую цепь. Следовательно, если калорийность растительного организма 1000 Дж, при полном поедании его травоядным животным в теле последнего останется из этой порции всего 100, в теле хищника – лишь 10 Дж, а если этот хищник будет съеден другим, то на его долю придется только 1 Дж, т. е. 0,1 %. Следовательно, запас энергии, накопленный зелеными растениями, в цепях питания стремительно иссякает. Поэтому пищевая цепь включает обычно всего 4– 5 звеньев. Потерянная в цепях питания энергия может быть восполнена только поступлением новых ее порций. Поэтому в экосистемах не может быть круговорота энергии, аналогичного круговороту веществ (рис. 147). Экосистема функционирует только за счет направленного потока энергии, постоянного поступления ее извне в виде солнечного излучения или готовых запасов органического вещества. 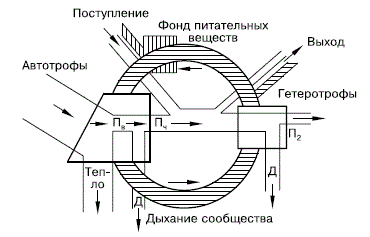
Рисунок 147 –Схема биогеохимического круговорота на фоне потока энергии (по Ю. Одуму, 1975): Пв – валовая продукция; Пч – чистая продукция; П2 – вторичная продукция; Д – траты энергии на обмен веществ (траты на дыхание); заштрихованная часть рисунка – круговорот вещества
Эффективность использования. Если в экосистеме не происходит постоянного накопления органического материала, то эффективность использования на каждом трофическом уровне, включая детритоядных, отражает всю чистую продукцию нижележащего трофического уровня. Исключение из этого правила составляют торфяные болота: значительная доля продукции растений опускается на дно болота, где кислые анаэробные условия препятствуют ее распаду. Однако большая часть экосистем характеризуется стационарными условиями, в которых вся продукция, в конечном счете, поглащается или выносится из системы ветром или водой. Поток энергии от одного трофического уровня к другому и так далее представляет собой сумму потоков, связанных с активностью питания многих видов. Отдельные популяции обычно потребляют лишь небольшую часть наличных пищевых ресурсов. Наибелее эффективны в этом отношении плотоядные, семеноядные и водные растительноядные; отдельные виды потребляют от 10 до 100% доступной им пищи. Наземные растительноядные обычно поедают лишь 1– 10% зеленой растительности. Охота за разными видами жертвы происходит с различным успехом. Так, исследование, проведенное в провинции Альберта (Канада), показало, что краснохвостые канюки летом вылавливают от 20 до 60% локальной популяции сусликов, но только 1% американских зайцев– беляков и 1– 3% воротничковых рябчиков. Другие хищники – лисы, куницы, совы, змеи – тоже охотятся на эти же самые виды жертв и обеспечивают перенос их биомассы на следующий трофический уровень. Те особи растений и жертвы, которые избежали уничтожения растительноядными и хищниками, в конце концов гибнут, и их химическая энергия поступает на следующий трофический уровень по детритному пути. Неудача одного хищника означает удачу другого, однако относительная эффективность питания консументов оказывает влияние на два важных аспекта энергетики сообщества: на количество энергии, которое передается в экосистеме по детритному пути, и на продолжительность времени, в течении которого энергия находится в данной системе, прежде чем она рассеивается в форме тепла. Детритный путь в экосистеме. Детритоядные виды поглощают остатки мертвых растений и животных, непереваренный или частично переваренный материал, выделяемый в виде экскрементов, и выделяемые азотсодержащие конечные продукты метаболизма белков – иными словами, любой неживой органический материал, который можно метаболизировать для извлечения энергии. Питание детритоядных организмов находится в обратной зависимости от перевариваемости свежей пищи. В планктонных водных сообществах детритоядные организмы играют менее важную роль, чем в наземных сообществах, где они потребляют до 90– 95% всей чистой первичной продукции. Обычно в качестве детритоядных членов естественных сообществ внимание привлекают крупные падальщики, как, например, грифы и вороны на суше и крабы в море, однако большую часть мервтого органического вещества поглощают малозаметные организмы, такие, как черви, клещи, бактерии и грибы. Из всех сообществ, основанных на детрите, наиболее хорошо изучены организмы, потребляющие лиственный опад и ветки, составляющие лесную подстилку. В широколиственном лесу растительноядные потребляют менее 10% продукции, если не считать нашестивия таких насекомых, как непарный шелкопряд, уничтожающий всю листву. Остальная продукция опадает на землю в виде сухих листьев и веток или же накапливается в корнях, стволах и ветвях, где она недоступна для консументов до тех пор, пока дерево в конце концов не отомрет. Разложение лиственного опада происходит тремя способами: 1) вымывание растворимых минеральных веществ и малых органических молекул водой; 2) потребление листьев крупными детритоядными организмами (многоножки, мокрицы и другие беспозвоночные); 3) окончательное разложение органических соединений до неорганических специализированными бактериями и грибами. От 10 до 30% веществ, содержавшихся в свежеопавших листьях, растворяются в холодной воде; большая часть этих веществ быстро удаляется из подстилки вымыванием. При дальнейшем разложении подстилки микроорганизмами образуются много малых органических и неорганических молекул, которые также вымываются, если их не успели ассимилировать детритоядные организмы. Крупные детритоядные ассимлируют не более 30– 45% энергии, заключенной в лиственном опаде, и еще менее того – из древесины. Тем не менее они ускоряют разложение подстилки микроорганизмами, перемалывая листья в своих пищеварительных трактах, размельчая подстилку и создавая новые поверхности для питания микроорганизмов. Листья деревьев разных видов разлагаются с различной скоростью, зависящей от их состава. Например, в восточной части штата Теннеси (США) потеря веса листьями в течение первого года после их опадения варьирует от 64% у шелковицы до 39% у дуба, 32% сахарного клена и 21% у бука. Иглы сосны и других хвойных также разлагаются медленно. Эти различия между видами в значительной степени зависят от содержания лигнина в их листьях. Лигнин – вещество, придающее дереву многие из его структурных качеств, и оно переваривается даже с большим трудом, чем целлюлоза. Иглы хвойных деревьев обычно содержат 20– 30% лигнина, а листья широколиственных видов – 15– 20%. В связи с устойчивостью некоторых типов растительной подстилки, особенно древесины, становится ясной роль грибов как детритоядных организмов. У большинства грибов имеется мицелий – обширная сеть нитевидных гиф, способных проникать в деревянистые клетки тех частей растительной подстилки, куда не могут проникнуть бактерии. Кроме того, грибы вырабатывают ферменты и кислоты, выделяя их непосредственно в субстрат, и тем самым способны переваривать органическое вещество даже на расстоянии. Грибы особенно многочисленны в лесной подстилке, с которой трудно справляться более крупным детритоядным организмам, а в тех местах, где дождевые черви и крупные членистоногие размельчают детрит, более многочисленны бактерии. Временная шкала потока энергии Растения обладают рядом качеств, препятствующих их перевариванию животными, что замедляет прохождение ассимилированной энергии по экосистеме. Для данного уровня продуктивности транзитное время, т.е. время, затрачиваемое на перенос энергии в данном сообществе, и запасание химической энергии в живой биомассе и детрите непосредственно связаны между собой: чем больше время переноса, тем больше накапливается живой и мертвой биомассы. Запасание энергии в экосистеме имеет большое значение для стабильности биологического сообщества и его способности противостоять разного рода воздействиям. Среднее время переноса энергии в живых организмах, входящих в состав данного сообщества, равно энергии, запасенной в этой системе в виде биомассы, деленной на скорость потока энергии через систему, или Время переноса (годы)= Биомасса (г/м2) / Чистая продуктивность(г/[м2.год]). Время переноса, определяемое этим отношением, иногда называют коэффициентом накопления биомассы. Значения среднего времени переноса для ряда экосистем варьируб от более чем 20 лет для лесных наземных экосистем до менее 20 дней в водных планктонных сообществах (табл. 13). Эти цифры представляют собой, однако, недооценку общего времени переноса энергии в данной экосистеме, потому что в них не учитывается накопление в подстилке мертвого органического вещества. Таблица 13 – Среднее время переноса энергии в живой растительной биомассе для некоторых экосистем (по Р. Риклефсу, 1979)
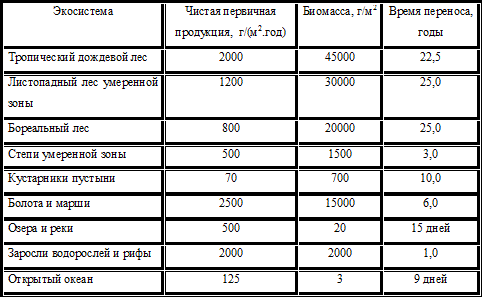
Оценку времени переноса или времени пребывания энергии в накопленной подстилке можно получить при помощи выражения, аналогичного предыдущему: Время переноса (годы)= Накопление подстилки (г/м2) / Скорость опада в подстилку (г/[м2.год]). Среднее время переноса для энергии варьирует от минимального, равного 3 месяцам, во влажных тропических лесах, до 1– 2 лет в сухих и горных тропических лесах, 4– 16 лет в сосновых лесах на юго– востоке США и до более чем 100 лет в хвойных лесах, растущих на горах. Высокие температуры и обилие влаги в низменных тропических лесах создают оптимальные условия для быстрого разложения. Ввиду того что большая часть энергии рассеивается детритоядными организмами, для получения полной оценки времени пребывания ассимилированной энергии в экосистеме среднее время переноса энергии следует добавить к времени ее переноса в живой растительности. Более прямые оценки скорости потока энергии можно получить, используя радиоактивные метки. При этом растения (или воду – при изучении водной системы) метят каким– нибудь радиоактивным изотопом, обычно 32Р, используемым в виде раствора фосфата. Затем через определенные промежутки времени после введения метки берутся выборки вида– консумента и исследуются с помощью счетчика излучения.
|