Тема 5.1. Понятие сукцессии как развития экосистемы
Любой биоценоз динамичен, в нем постоянно происходит изменение в состоянии и жизнедеятельности его членов и соотношении популяций. Все многообразные изменения, происходящие в любом сообществе, можно отнести к двум основным типам: циклические и поступательные. Циклические изменения сообществ отражают суточную, сезонную и многолетнюю периодичность внешних условий и проявления эндогенных ритмов организмов. Суточные циклы связаны главным образом с ритмикой природных явлений и носят строго периодический характер. Суточная динамика в биоценозах, как правило, выражена тем сильнее, чем значительнее разница температур, влажности и других факторов среды днем и ночью. В каждом биоценозе имеются группы организмов, активность жизни, у которых приходится на разное время суток. Одни активны днем, другие – ночью. Так, в песчаных пустынях Средней Азии жизнь летом в полуденные часы замирает. Даже виды с дневной активностью прячутся от жары в норах, в тени саксаула либо на его ветвях (агамы, ящерки). Ночью пустыня оживает. Ночных и сумеречных животных здесь больше, чем дневных. Многие дневные виды летом переходят к ночному образу жизни (большинство змей, пауки, чернотелки). Активно действуют жуки–копрофаги, вылетают козодои, домовые сычи, кормятся тушканчики, гекконы, хищничают фаланги, скорпионы, лисицы, корсаки, змеи. Суточные ритмы прослеживаются в сообществах всех зон, от тропиков до тундр. Даже при непрерывном освещении летом в тундре отмечаются суточные ритмы в распускании цветков у растений, питании птиц, в лёте и распределении насекомых и т. п. Сезонная изменчивость биоценозов выражается в изменении не только состояния и активности, но и количественного соотношения отдельных видов в зависимости от циклов их размножения, сезонных миграций, отмирания отдельных генераций в течение года и т. п. На определенное время года многие виды практически полностью выключаются из жизни сообщества, переходя в состояние глубокого покоя (оцепенения, спячки, диапаузы), переживая неблагоприятный период на стадии яиц и семян, перекочевывая или улетая в другие биотопы или географические районы. Сезонной изменчивости подвержена зачастую и ярусная структура биоценоза: отдельные ярусы растений могут полностью исчезать в соответствующие сезоны года, например, травянистый ярус, состоящий из однолетников. Сезонные ритмы сообществ наиболее отчетливо выражены в климатических зонах и областях с контрастными условиями лета и зимы. В слабой форме они, однако, прослеживаются даже в тропических дождевых лесах, где длительность дня, температура и режим влажности очень мало меняются в течение года. Многолетняя изменчивость– нормальное явление в жизни любого биоценоза. Она зависит от изменения по годам метеорологических условий (климатических флюктуации) или других внешних факторов, действующих на сообщество (например, степени разлива рек). Кроме того, многолетняя периодичность может быть связана с особенностями жизненного цикла растений–эдификаторов, с повторением массовых размножений животных или патогенных для растений микроорганизмов и т. д. Примером могут служить изменения, происходящие на лугах лесостепной Барабы. Количество осадков, выпадающих в Барабинской лесостепи, резко колеблется по годам, и, как правило, ряд засушливых лет чередуется с многолетним периодом обилия осадков. Постепенное понижение уровня почвенно–грунтовых вод в засушливый период 1950–1955 гг. стимулировало проникновение вслед за ними корневых систем глубокоукореняющихся трав, таких, как колосняк мохнатый, кермек Гмелина и др. У этих видов отсутствовала реакция на иссушение почвы, но при дальнейшем повышении уровня грунтовых вод они пострадали в первую очередь из–за отмирания затопленных водой корневых окончаний. Засуха особенно сильно сказалась на мезофильных мелкоукореняющихся видах – овсянице красной, полевице тонкой, лапчатке гусиной и других, а также на гидрофитах – ситнике, вейнике и пр. Они или гибли, изреживались, или переходили в состояние покоя. Усиливавшееся в засуху засоление почв послужило причиной массовой гибели от солевого отравления овсяницы красной. Во влажные годы, когда промачивание почвы привело к ее опреснению в верхних слоях, этот вид появился вновь. Высокие температуры и усиление аэрации почвы способствовали развитию бактерий–нитрификаторов и накоплению нитратов. Это стимулировало массовое размножение в разнотравье осота полевого, одуванчика бессарабского и некоторых других азотолюбивых видов. Когда энергия нитрификации вслед за увеличением влажности почв снизилась, эти виды, не выдержав конкуренции со злаками, перешли в угнетенное состояние. Изменения в фитоценозах были усилены при массовом размножении в 1953 г. нестадных саранчовых. Прямокрылые съели до 80–90 % массы мягколистных злаков: пырея, костра, типчака и др. У жестколистных колосняка, тростника и других они уничтожали всходы по мере их появления. В этот год бурно разрослось непоедаемое саранчовыми разнотравье, для которого была устранена конкуренция со стороны злаков. Последствия размножения саранчовых сказались и на соотношении злаков в последующие два года. В засушливые годы муравьи Lasius niger и Myrmica scabrinodis заселили, кроме возвышенностей, высохшие низинные луга, где начали земляную строительную деятельность, которая повлияла и на условия произрастания растений. Подобные многолетние изменения в составе ценозов имеют тенденцию повторяться вслед за периодическими локальными изменениями климата, которые связаны с изменением общей циркуляции атмосферы, обусловленной, в свою очередь, усилением и ослаблением солнечной активности. Вследствие многостепенной зависимости изменения в сообществах не прямо отражают ритмику космических процессов. Примером обратимых изменений фитоценозов, связанных с особенностями жизненного цикла растений–эдификаторов, могут быть преобразования в буковом лесу. Взрослые буки с плотно сомкнутыми кронами создают такую густую тень, что в лесу практически отсутствуют подлесок и травяной покров, а сеянцы буков пребывают в угнетенном состоянии и погибают. Когда старые деревья достигают предельного возраста и падают, через появившиеся «окна» на почву проникает солнечный свет, и трогаются в рост молодые буки. Некоторое время древостой характеризуется разновозрастностью, пока не выпадут все старые деревья и не выйдут в первый ярус подросшие молодые буки, после чего вновь устанавливается практически одноярусная структура фитоценоза. Весь цикл занимает около двух с половиной столетий. Примером многолетних изменений экосистем, вызываемых биотическими причинами, является динамика степных экосистем Монголии под влиянием вспышек численности полевки Брандта – мышевидного грызуна, который является «ключевым» видом. При массовом развитии полевки резко меняется состав растительного сообщества: вместо ковылей, листья которых съедают грызуны, из подземных корневищ развиваются побеги других злаков, особенно востреца (Elymus chinensis). Однако вслед за пиком численности начинается спад плотности популяции грызуна. И через несколько лет популяции ковылей также восстанавливаются, а корневищные злаки переходят в прежнее состояние «полупокоя» и «готовятся» к новой вспышке обилия грызунов. Колебания фаз «Elymus chinensis – Stipa krilovii» – характерная особенность монгольских степей, которую описали выдающиеся исследователи А.А. Юнатов и Е.М. Лавренко. В европейских широколиственных лесах в некоторые годы массово развивается непарный шелкопряд. Его гусеницы почти полностью поедают листву деревьев, что улучшает условия для жизни растений напочвенного покрова (освещенность, обеспеченность элементами минерального питания за счет экскрементов гусениц). В итоге резко падает биологическая продукция деревьев, но возрастает продукция трав и соответственно связанных с ними фитофагов. Кабаны постоянно перерывают участки леса в поисках корма. На пороях площадью несколько десятков метров разрастаются рудеральные растения, однако в течение 4–5 лет происходит восстановление напочвенного покрова и как следствие – циклическая динамика всей биоты. Естественно, что «вспашка» кабанами участка леса резко изменяет жизнь всего почвенного ценоза. Активизируется деятельность бактерий–аэробов и животных, предпочитающих условия рыхлых и хорошо аэрированных почв. Более продолжительны циклы, вызываемые деятельностью бобров: после того, как они запрудят реку, в течение нескольких лет происходит интенсивная перестройка экосистемы и возрастает роль влаголюбивых растений и их спутников. Виды деревьев, неустойчивых к подтоплению и затоплению, вообще погибают. Однако за 10–20 лет использования этой территории бобры выедают растения, служащие им кормовой базой (в первую очередь ольху) и меняют место жительства. Происходит достаточно быстрое разрушение «гидромелиорированной» экосистемы и восстановление прежней. Этот цикл продолжается примерно 100 лет. В масштабе десятилетий происходят обратимые изменения лесов Дальнего Востока, связанные с биологическими циклами видов бамбука из рода Sasa, являющихся ключевыми в этих экосистемах. Бамбуки, развивающиеся в подлеске, подавляют возобновление деревьев. Но они монокрапики (т.е. плодоносят всего один раз и после этого погибают), и после гибели очередной генерации бамбука в течение нескольких лет до следующего его разрастания активно возобновляются популяции деревьев. В широколиственных лесах Восточной Европы в результате выпадения отдельных деревьев (от старости или под влиянием ветра) образуются «окна». В «окнах» размером несколько десятков метров формируются сообщества из эксплерентов (рудеральных трав, ольхи, березы), которые спустя несколько десятилетий замещаются «основными» видами этого типа леса. Исследователи тропических лесов назвали эти сменяющие друг друга группы «дриадами» и «номадами». Динамика «дриад» и «номад» соответствует одной из моделей устойчивости экосистем: устойчивость в крупном масштабе биологического пространства слагается из неустойчивостей в его мелком масштабе. В целом любые циклические изменения экосистем – это отражение их пластичности, т.е. приспособленности состава, структуры и функций к колебаниям условий среды и жизненным циклам «ключевых» видов. Таким образом, многолетняя цикличность зависит от изменения по годам метеорологических условий (климатических флюктуации), неравномерным выпадением осадков по годам, с периодическим повторением засух или других внешних факторов, действующих на сообщество (например, степени разлива рек). Поступательные изменения в сообществе приводят в конечном итоге к смене этого сообщества другим, с иным набором господствующих видов. Причиной подобных смен могут быть внешние по отношению к биоценозу факторы, длительное время действующие в одном направлении, например возрастающее в результате мелиорации иссушение болотных почв, увеличивающееся загрязнение водоемов, усиленный выпас скота, вытаптывание лесопарков населением городов и т. п. Возникающие при этом смены одного биоценоза другим называют экзогенетическими. Если при этом усиливающееся влияние фактора приводит к постепенному упрощению структуры сообществ, обеднению их состава, снижению продуктивности, то подобные смены называют дигрессионными или дигрессиями. Так, пастбищные дигрессии на террасовых песках Нижнего Днепра развиваются следующим образом. При умеренном выпасе степь находится на стадии дерновинных злаков. Преобладают типчак, кипец, житняк, ковыль. При дальнейшей усиленной пастьбе возникает стадия стержнекорневых двудольных с господством чаще всего молочая, а также двулетников и однолетников. Дерновинки злаков разбиваются копытами скота, а затем почти совершенно исчезают. На третьей стадии корневищных растений появляются злаки, характерные для сыпучих и слабозаросших песков: песчаный пырей, вейник, осока песчаная. На следующей стадии возникают голые пески с отдельными зарослями псаммофитов, характерных для предыдущего этапа. Таким образом, ковыльно–типчаковая степь сменяется сыпучими песками. Соответственно меняется и характер животного населения. Эндогенетические смены возникают в результате процессов, происходящих внутри самого сообщества. Сукцессия (от лат. сукцессио – последовательность, смена) – последовательная смена биоценозов, преемственно возникающая на одной и той же территории (биотопе) под влиянием природных факторов или воздействия человека. В основе сукцессии лежит неполнота биологического круговорота в данном ценозе. Каждый живой организм в результате жизнедеятельности меняет вокруг себя среду, изымая из нее часть веществ и насыщая ее продуктами метаболизма. При более или менее длительном существовании популяций они меняют свое окружение в неблагоприятную сторону и в результате оказываются вытесненными популяциями других видов, для которых вызванные преобразования среды оказываются экологически выгодными. Таким образом, в сообществе происходит смена господствующих видов. Длительное существование биоценоза возможно лишь в том случае, если изменения среды, вызванные деятельностью одних организмов, точно компенсируются деятельностью других, с противоположными экологическими требованиями. В ходе сукцессии на основе конкурентных взаимодействий видов происходит постепенное формирование более устойчивых комбинаций, соответствующих конкретным абиотическим условиям среды. Примерами сукцессий могут быть смены видов при зарастании стоячих водоемов (рис. 155) или барханных песков в пустыне. 
Рисунок 155 – Изменение экосистемы в ходе сукцессии при зарастании озера (по Д. Казенс, 1982): 1–6– смена доминирующих видов растений
Различают ненаправленные и направленные смены растительного покрова. Ненаправленные смены представляют собой дальнейшее углубление процесса погодичных (разногодичных) изменений в пределах фитоценоза. Направленные смены представляют собой необратимые изменения растительного покрова, при которых одни сообщества сменяются другими. И это уже сукцессии. В России изучение сукцессий лесов было начато Г.Ф. Морозовым и продолжено В.Н. Сукачевым, Е.М. Лавренко, П.Д. Ярошенко и другими лесоводами, а исследование смен растительности на степных залежах и выгонах – И.К. Пачоским, Г.Н. Высоцким, Л.Г. Раменским и др. За рубежом особенно детально изучали сукцессии американские ученые, в первую очередь Ф. Клементс и Д. Уивер, английские ботаники во главе с А. Тенсли, ученые франко–швейцарской школы, руководимые Ж. Браун–Бланке. Сукцессия растительности – это последовательный ряд смены серийных (временно существующих) растительных сообществ на конкретном местообитании после выведения конкретной экосистемы из состояния динамического равновесия. При этом конкретная экосистема возвращается в свое исходное состояние и пребывает в нем до тех пор, пока не изменятся климат, рельеф, гидрологический режим, пока вновь не пройдет пожар, или не случится какая–то другая катастрофа. И вновь начнется новая сукцессия, которая либо приведет к восстановлению исходного сообщества, либо нет. В результате сукцессии на конкретном местообитании восстанавливается исходное растительное сообщество, или коренное. Коренное сообщество растений устойчиво и в данных климатических условиях не изменяется. Необратимость процесса – обязательное свойство любой сукцессии. Смены растительности на вырубках или на залежах, приводящие к восстановлению коренной растительности, считают сукцессиями. Всю конкретную сукцессию в целом называют серией (американские авторы). Каждое сообщество, которое в процессе сукцессии сменяет другое, называют стадией этой сукцессии, или серийным сообществом. Заключительное сообщество, находящееся в относительном соответствии с условиями существования, называют климаксом (американские авторы) или узловым сообществом (Ярошенко). В одном и том же месте однотипная серия может повторяться неоднократно, если повторяются нарушения растительного покрова или его уничтожение, или резко меняются условия существования. Сыпучие барханные пески Каракумов и других районов Средней Азии сначала полностью лишены растительности и постоянного населения животных. Первым на них поселяется многолетний злак аристида, хорошо приспособленный к жизни в условиях постоянного переноса песка ветром. Корни у этого растения шнуровидные, и каждый заключен в чехол из сцементированных песчинок, что защищает корни от высыхания и механического повреждения, если они окажутся на поверхности. Побеги растут вертикально вверх и образуют дополнительные зоны кущения выше материнского, если песок засыпает растение. За счет аристиды уже могут существовать некоторые насекомые, и поэтому на барханы начинают забегать в поисках пищи ящерицы рода Eremias. На слегка скрепленных корнями аристиды песках получает возможность поселиться длиннокорневищная песчаная осока. Она успешно борется с песком, быстро прорастая сквозь его наносы и пронизывая песок ветвящимися корневищами на глубине 1–5 см. Покров разрежен, так как для обеспечения влагой одного растения нужна значительная площадь. На скрепленных злаками и осокой песках поселяются затем кустарники джузгун и белый саксаул, а также другие травянистые растения, в основном эфемеры: злаки, крестоцветные, мотыльковые, бурачниковые и т. п. Вслед за растительностью появляются растительноядные млекопитающие: тонкопалый суслик, мохноногий тушканчик, полуденная песчанка. Увеличивается видовое разнообразие насекомых – кормовой базы ящериц: ушастой и песчаной круглоголовок, сетчатой ящурки, гекконов. Появляются птицы – саксаульная сойка, дрофа–красотка, затем змея и хищные млекопитающие. Закрепленные пески Средней Азии отличаются большим видовым богатством и разнообразием жизненных форм растений и животных, так как водный режим их достаточно благоприятен: пески обладают способностью сгущать водяные пары в почве во влагу. Сукцессии в природе чрезвычайно разномасштабны. Их можно наблюдать в пробирках с сенным настоем, где сменяют друг друга разные виды микроорганизмов и простейших (рис. 156), в лужах и прудах, в одной и той же порции листового опада в лесу, где происходят закономерные смены сапрофитов и сапрофагов, на стволах отмирающих деревьев, в пнях, на кротовинах, выбросах из нор сурков, зарастающих отмелях, выветривающихся скалах, на заброшенных пашнях, насыпях, на болотах, лугах, в лесах и т. д. Иерархичность в организации сообществ проявляется и в иерархичности сукцессионных процессов: более крупные преобразования биоценозов складываются из более мелких. Даже в стабильных экосистемах с хорошо отрегулированным круговоротом веществ постоянно осуществляется множество локальных сукцессионных смен, поддерживающих сложную внутреннюю структуру сообществ. Изменения в сообществе в результате сукцессии носят закономерный характер и обусловлены взаимодействием организмов между собой и окружающей абиотической средой. Экологическая сукцессия происходит в определенный отрезок времени, в который изменяется видовая структура сообщества и абиотическая среда его существования вплоть до кульминации его развития – возникновения стабилизированной системы. В этом состоянии система находится тогда, когда в ней на единицу энергии приходится максимальная биомасса и максимальное количество симбиотических связей между организмами. 
Рисунок 156 – Смена простейших в сенном настое (по Р. Дажо, 1975):
А – общая схема; Б – конкретная серия: 1 – жгутиковые; 2 – инфузории кольноды; 3 – нарамеции; 4 – брюхо–ресничные инфузории; 5 – амебы; 6 – сувойки
Типы сукцессионных смен. Выделяют два основных типа сукцессионных смен: 1) с участием как автотрофного, так и гетеротрофного населения и 2) с участием лишь гетеротрофов. Сукцессии второго типа совершаются только в таких условиях, где создается предварительный запас или постоянное поступление органических соединений, за счет которых существует сообщество: в сильно загрязненных органическими веществами водоемах, в скоплениях разлагающейся растительной массы, в кучках или буртах навоза, компостах, в пещерах с гуано летучих мышей и т. п. Сукцессии со сменой растительности могут быть первичными и вторичными. Первичная сукцессия начинается на субстрате, практически не измененном деятельностью организмов (на скалах, песках, обрывах). Вторичная сукцессия– это последовательная смена одного сообщества, существовавшего на данном субстрате, другим более совершенным для данных абиотических процессов. Первичная сукцессия позволяет проследить формирование сообществ с самого начала (на склоне после оползня или обвала, на образовавшейся отмели при отступлении моря, свежая лесосека, искусственные водохранилища). Классический пример природной сукцессии – «старение» озерных экосистем – эвтрофикация. Она выражается в зарастании озер растениями от берегов к центру. Здесь наблюдается ряд стадий зарастания. В конечном итоге озеро превращается в торфяное болото, представляющее устойчивую экосистему климаксного типа. Такие сукцессии в геоботанике называют экогенетическими, так как они ведут к преобразованию самого местообитания. Первичные автогенные сукцессии зарастания субстратов, образующихся после таяния ледника на Новой Земле, еще в начале XIX столетия описал русский ученый К. Бэр (Трасс, 1976). Тем не менее концепция первичной автогенной сукцессии, в результате которой экосистема переходит в экологически равновесное состояние, наиболее соответствующее климату, связана с именем выдающегося американского эколога Ф. Клементса. Это равновесное состояние было названо климаксом. Экосистемы стадий сукцессии на пути к климаксу Клементс назвал серийными. Клементс считал, что в любом географическом районе с одним типом климата есть только один тип экосистемы (моноклимакс), который наиболее соответствует этому климату. К примеру, в Восточной Европе в биоме тайги – это еловый лес, в биоме широколиственных лесов – липово–дубовый лес, в биоме степей – разнотравно–ковыльная степь. Все иные типы экосистем «стремятся» перейти в этот тип, т.е. происходит процесс конвергенции (выравнивания) состава экосистем одного района: на скалах образуются почвы; озера зарастают, превращаясь в болота, которые со временем высыхают; происходит измельчение минеральных частиц (пески превращаются в суглинки); более сухие местообитания становятся более влажными за счет накопления органического вещества, которое способно удерживать дождевые и снеговые воды. Кроме того, Клементс выделял множество разных типов сообществ (и соответствующих им экосистем), которые в результате действия какого–либо внешнего фактора «застревают» на определенной стадии сукцессии и не могут перейти в климакс, т.е. являются хронически сериальными. Например, субклимакс – это экосистема поймы реки, которая не переходит в климакс вследствие регулярных паводков. Дисклимакс – это экосистема, которая не переходит в климакс в результате действия нарушающего ее фактора (например интенсивно использующееся пастбище). В ходе сукцессий экосистем, формирующих климакс, возрастают продуктивность и биомасса, видовое богатство, сложность структуры (формируются почвы, появляются растения разных жизненных форм – деревья, кустарники, травы, что формирует дополнительные ниши для гетеротрофов). Повышается роль различных механизмов сосуществования – дифференциации экологических ниш, мутуализма, коадаптации между хищниками и их жертвами и т.д. Условия для жизни растений и видов гетеротрофной биоты в ходе такой сукцессии улучшаются, а сама последовательность видов в ходе сукцессии жестко детерминирована законами «онтогенеза» экосистем. Экологи, развивавшие функциональный взгляд на экосистему (А. Лотка, Г. Одум, Р. Пинкертон, Р. Маргалеф) подчеркивали, что по мере приближения к климаксу происходит сдвиг потока энергии от продуктивности к дыханию (Лотка говорил даже о «законе максимума биологической энергии»). Ю. Одум (1986) подчеркнул, что в ходе сукцессии по мере ее приближения к климаксу происходит выравнивание соотношения продукции (P) и дыхания (R), т.е. в климаксовой экосистеме Р=R. Вся продукция, которая образована за год, растрачивается на дыхание, и потому дальнейшего увеличения биомассы не происходит. Отношение величины биомассы к продукции (В/Р) возрастает до тех пор, пока на единицу потока энергии не будет приходится максимум биомассы для данного климата (этот максимум будет различаться в зонах тайги, широколиственных лесов, степи, пустыни и т.д.). По мере приближения к климаксу круговороты биогенных элементов становятся все более замкнутыми и медленными, причем возрастает доля биогенов, которые фиксированы в живых организмах и детрите (включая и гумус почвы). В ходе сукцессии происходят «эстафеты» представителей флоры, фауны, грибов, микроорганизмов, причем в большинстве случаев виды r–стратегии сменяются видами К–стратегии (по Макартуру и Уилсону) или (по Раменскому и Грайму) виды стратегии R – видами стратегий С, S и различных переходных вторичных типов (CS, CR, RS, CRS). Таким образом, малолетники сменяются многолетниками, а травы – деревьями, что приводит к увеличению биологической продукции за счет более полного использования ресурсов. Работы Клементса навсегда останутся классикой экологии и краеугольным камнем теории динамики экосистем. Тем не менее сформулированные им представления в ходе дальнейшего развития экологии претерпели существенные изменения: 1. А. Тенсли и А. Найколсон показали, что в одном районе может формироваться не один, а несколько климаксов, т.е. экосистемы, которые формируются при сукцессиях зарастания скал, озер, песков, лессовидных суглинков и т.д., будут различными. Концепция моноклимакса, таким образом, переросла в концепцию поликлимакса. Р. Уиттекер, развивая эти представления, сформулировал концепцию «климакс–континуума». Он считал, что разные экосистемы поликлимакса связаны друг с другом плавными переходами и по этой причине в каждой точке – свой климакс. 2. Климакс – это не обязательно самая продуктивная и богатая видами экосистема. Как правило, наибольшим видовым богатством и продуктивностью отличаются как раз «предклимаксовые» серийные экосистемы. 3. Сукцессия не является жестко детерминированным, «запрограмированным» процессом, подобным онтогенезу организма, а имеет стохастический характер. Закономерности сукцессий можно выявить только при обобщении (усреднении) результатов наблюдений за несколькими конкретными сукцессиями, протекающими в одних и тех же условиях. В конкретных сукцессионных последовательностях приход видов в сукцессию и уход из нее может происходить в разной очередности. Более того, некоторые виды могут участвовать в одной конкретной сукцессии и не участвовать в другой. Ф. Клементс считал, что все сукцессии развития экосистем в направлении климакса подчиняются одной модели: улучшаются условия для жизни биоты, и потому возрастают биологическая продукция и видовое богатство экосистемы. Современные экологи различают не менее трех моделей сукцессий (Connell, Slayter, 1977): – модель благоприятствования. Соответствует представлениям о сукцессии Клементса: продуктивность и видовое богатство в ходе сукцессии возрастают вплоть до стадии климакса. Классический пример такой сукцессии – зарастание скал, где последовательно сменяют друг друга стадии цианобактерий и водорослей, накипных лишайников, кустистых лишайников и мхов, трав, кустарников и деревьев; – модель толерантности. В ходе сукцессии условия ухудшаются, пример – переход низинного болота в верховое, при котором происходит ухудшение условий минерального питания, и потому снижаются продуктивность и видовое богатство. Ухудшаются условия для жизни биоты и в ходе сукцессии на богатых субстратах: первым растениям–поселенцам достается больше ресурсов минерального питания и света, чем вторым и третьим, которые должны обеспечивать себя ресурсами в условиях возрастающей конкуренции; – модель ингибирования. В ходе сукцессии появляется «ключевой» вид (или гильдия ключевых видов), который блокирует дальнейшие изменения. В результате происходит остановка сукцессии и она не доходит до стадии климакса. Например на лесных гарях в Шотландии кукушкин лен блокирует поселение деревьев, в пустынях Средней Азии поселению кустарников и саксаула препятствует корка, которую образуют цианобактерии, водоросли и некоторые мхи. Восстановление прерии в Северной Америке блокируется разрастанием заносных европейских злаков–однолетников, в первую очередь Bromus tectorum. В ходе сукцессии может происходить смена модели благоприятствования моделью толерантности: на первых стадиях условия улучшаются, а по мере приближения к климаксу – ухудшаются. Наглядный пример сукцессии со сменой модели –формирование растительности при освобождении побережья фиорда ото льда на Аляске (Chapin et al., 1994). Выделяется четыре стадии процесса: – пионерная (до 20 лет). Поверхность субстрата покрывается «черной коркой» из азотфиксирующих цианобактерий, гаметофитов хвоща (Equisetum variegatum), лишайников, печеночников, на фоне которых рассеянно встречаются травы, кустарничек Dryas drummondii, отдельные экземпляры ивы, тополя (Populus trichocarpa), ели (Picea sitchensis) и ольхи (Alnus sinuata); – стадия Dryas (между 20 и 30 годами). Вся поверхность покрыта ковром кустарника, в котором рассеяны одиночные экземпляры ив, тополей, елей и ольхи; – стадия ольхи (между 50 и 100 годами); – стадия ели (после 100 лет). В ходе сукцессии формируется почва, которая обогащается органическим веществом и азотом, а смена видов растений идет в направлении повышения их высоты и длительности жизни, что соответствует модели благоприятствования. Однако при этом возрастает уровень конкуренции за свет и почвенные ресурсы (особенно на стадии ели), ухудшаются условия для приживания всходов, повышается вероятность гибели семян, что соответствует модели толерантности. смена моделей происходит на четвертой стадии. Смена модели благоприятствования моделью толерантности характерна и для сукцессий экосистем в теплом климате. Так при зарастании лавовых потоков на первых стадиях условия улучшаются за счет бобовых (особенно из р. Lupinus), которые способствуют обогащению субстрата азотом, а в дальнейшем – ухудшаются, так как обостряется конкуренция. Несмотря на то, что автогенные сукцессии протекают спонтанно по присущим им внутренним законам, человек, зная эти законы, может влиять на скорость сукцессии. Так для ускорения самозарастания отвалов пустой породы их поверхность покрывается тонким слоем торфа или почвы, в которой содержатся семена растений. Кроме того, процесс зарастания может быть ускорен посевом семян луговых трав или посадкой кустарников и деревьев. В первичных сукцессиях, протекающих на скалах в лесах Урала, различают четыре этапа. 1. Поселение эндолитических и накипных лишайников, которые сплошь покрывают каменистую поверхность. Накипные лишайники несут своеобразную микрофлору и содержат богатую фауну простейших, коловраток и нематод. Мелкие клещи–сапрофаги и первичнобескрылые насекомые обнаруживаются сначала только в трещинах. Активность всего населения прерывиста, в основном после смачивания скал дождем или туманом. 2. Преобладание листоватых лишайников, постепенно образующих сплошной ковер. Под круговинками лишайников Parmelia в результате действия выделяемых ими кислот и механического сокращения слоевищ при высыхании образуются выщербленности, идет отмирание слоевищ и накопление детрита. Под лишайниками в большом количестве обитают мелкие членистоногие: коллемболы, панцирные клещи, сеноеды, личинки комаров–толкунчиков и другие, образуется микрогоризонт, состоящий из их экскрементов. 3. Поселения литофильных мхов Hedwigia ciliata и Pleurozium schreberi. Они погребают лишайники и подлишайниковые пленочные почвы. Ризоиды мхов прикрепляются не к камню, а к мелкозему, мощность которого достигает уже 3 см. Под мхами колебания температуры и влажности уже в несколько раз меньше, чем под лишайниками. Усиливается деятельность микроорганизмов, и увеличивается разнообразие групп животных. 4. Появление гипновых мхов и сосудистых растений. В разложении растительных остатков и формировании почвенного профиля постепенно уменьшается роль мелких членистоногих и растет участие более крупных беспозвоночных–сапрофагов: энхитреид, дождевых червей, личинок насекомых. Сукцессии могут начинаться на участках, полностью свободных от растительного покрова и не сохранивших семенных зачатков растений (первичные сукцессии). Образование участков, полностью лишенных растительности, и участков, где растительный покров в той или иной степени нарушен, вызывается действием ветра (ветровая эрозия или ветровая аккумуляция), отступлением берега водоема и заливанием участка водой, выгоранием растительного покрова, выгрызанием растений животными, вырубкой леса, распашкой земель, т. е. климатическими, эдафическими, биотическими и антропогенными факторами. Границу между флюктуационными изменениями (обратимыми сменами) в фитоценозе и сукцессиями (необратимыми сменами одного фитоценоза другим) провести трудно. Сукцессии накладываются на изменения, происходящие в растительном сообществе (суточные, сезонные, разногодичные, определяемые микроэволюцией видов, онтогенетические и возобновительные), а также на разногодичные смены растительных сообществ. Для того чтобы различить, каким изменениям соответствуют те или иные явления в жизни растительного покрова, необходимы стационарные исследования и сравнение фитоценозов, существующих в различных условиях и представляющих как бы проекцию в пространстве тех изменений, которые осуществляются в растительном покрове в течение определенного времени. А.Г. Воронов (1940, 1973) выделяет в процессе сукцессии растительности на обнаженных почвах или грунтах две фазы: – заселение оголенной территории и формирование фитоценоза из поселяющихся на голой площади растений – смена одного сформировавшегося фитоценоза другим. а) Факторы, определяющие развитие растительности в первой фазе сукцессии – на оголенных территориях Растения проникают на освободившуюся территорию путем переноса диаспор (семян, спор, кусочков растения) при помощи ветра, воды, животных или человека, или путем постепенного вегетативного разрастания астений, находящихся близ границ оголенной территории. Случаи внедрения растения на новую территорию с краев только путем образования вегетативных подземных или надземных побегов без образования генеративных органов наблюдаются во много раз реже, чем внедрение путем заноса диаспор. Успех переноса диаспор зависит от расстояния, отделяющего растение от заселяющейся территории, способа переноса диаспор, их количества, численности растений данного вида. При этом легкость семян, способствующая их переносу при помощи ветра, достигается за счет уменьшения запасов питательных веществ, а это отрицательно сказывается на развитии проростков, снижая шансы на их сохранение. Заселение новой территории зависит от ряда факторов, случайных по отношению к особенностям самой территории: – от того, какие растения растут поблизости от обнаженного участка, – от их количества, – от направления господствующего ветра, – от высоты и силы паводка, – от качества субстрата участков, – от характера увлажнения, и т. д. На первой стадии развития нового фитоценоза в его составе часто преобладают растения с диаспорами, легко разносимыми ветром, а около воды – с диаспорами, хорошо держащимися на воде. Часто то действие, которое вызвало оголение почвы (отложение наноса водой, перевевание песка ветром) способствует и появлению на этой территории диаспор. Поэтому так быстро вырастают сорные и рудеральные растения на участках, обнажение которых вызвано деятельностью человека. б) Эцезис и его особенности После того как растение проникло на оголенную территорию, оно начинает приспосабливаться к новым условиям. Этот процесс приспособления особей растений к новым для них условиям называют эцезисом. Он начинается с прорастания семян и заканчивается, когда растение дало плоды и семена. Не все диаспоры, попавшие на оголенную территорию, прорастают сразу. Семена большинства видов долго сохраняют всхожесть нередко десятки или даже сотни лет. При этом они прорастают не в один год, а при благоприятном стечении обстоятельств. Это обеспечивает условия лучшего сохранения проростков ПРИМЕР. Леспедеца в дубняках вокруг пос. Горнотаежеое (Уссурийский р–он) возобновилась в первый же год после пожара, образовав сплошной покров. Пожара до этого не было более 20 лет. Лишь немногие виды (конский каштан, чозения, ивы, и др.) имеют семена, теряющие всхожесть в течение нескольких дней или немногих недель. Семена, образующие почвенный запас, нередко принадлежат растениям разных жизненных форм и потому обеспечивают развитие растений при разнообразных условиях среды (семена одних видов прорастают при более высоких температурах, других – при более низких, одни при большей влажности грунта, другие – при меньшей и т. д). Внедрившиеся на оголенную территорию, растения начинают плодоносить и сами становятся источником диаспор. Теперь диаспоры попадают на заселяющуюся площадь не только со стороны, но и от тех растений, которые уже растут и плодоносят здесь же. На стадии проростков многие растения, развивающиеся на оголенных территориях, гибнут под действием неблагоприятных условий, в результате уничтожения животными или паразитическими растениями и вследствие конкуренции с другими проростками. В зависимости от условий существования оголенную территорию населяют один или несколько видов. Чем суровее условия, тем меньшее количество видов растений может начать здесь свое развитие. Наиболее бедный состав проростков типичен для сильно засоленных грунтов, обнажений скал и пр. Когда растение из стадии проростка переходит в более поздние стадии развития, его потребности в воде и пище возрастают, а запасы питательных веществ в семени или в плоде к этому времени исчерпываются, и растение целиком зависит от пищевых ресурсов внешней среды. Поэтому с ростом растений усиливается и конкуренция. В это время становится заметным и влияние самих растений (во всяком случае доминантов) на среду. При этом, чем суровее условия среды, тем большую роль для растений, внедряющихся на данную территорию, играет прямое влияние внешних условий и тем меньшее значение имеет конкуренция. Чем менее суровы условия среды, тем меньшую роль играют внешние условия и тем больше значение конкуренции. в) Стадии развития фитоценоза первичной сукцессии (по А.Г. Воронову) Пионерная группировка – случайное сочетание растений. Фитоценозы, формирующиеся на оголенных площадях, на первой стадии развития характеризуются: – случайным составом растений, – отсутствием сомкнутого растительного ковра, – слабым воздействием на среду и – почти полным отсутствием взаимовлияния между особями. Пионерная группировка может быть чистой (одновидовой), так и смешанной (многовидовой). Если условия среды быстро меняются в направлении возрастания суровости (например, грунт высыхает, засоляется и пр.), то число видов, поселившихся на техногенном участке, уменьшается, и смешанная пионерная группировка обедняется и в конце концов может перейти в чистую пионерную группировку. ПРИМЕР. На высохшем днище оз. Южная Сарпа (Западный Прикаспий) была обнаружена смешанная пионерная группировка, образованная марью красной (Chenopodium rubrum), верблюжьей колючкой (Hhagi camelorum), лебедой поникшеи (Atriplex patens), пыреем ползучим (Agropyron repens), скрытницей колючей (Crypsis aсuleata), турнефорцией сибирской (Tournefortia sibirica), вьюнком полевым (Convolvulus arvensis), канатником Феофраста (Аbutilon theophrasti). Как видно, экологические особенности видов, образующих смешанную пионерную группировку, весьма разнообразны. Простая группировка – следующая стадия развития фитоценоза после пионерной группировки. В такой группировке растительный покров: – в надземной части не сомкнут, однако растения располагаются значительно теснее, чем в пионерной группировке. – хорошо заметно взаимовлияние растений. – обычен групповой характер распределения растений: вокруг особи, давшей семена, развивается ее потомство. Простые группировки, как и пионерные, могут быть чистыми (одновидовыми) и смешанными (многовидовыми), образованными несколькими видами, и растения в них, в отличие от смешанных пионерных группировок, всегда относятся к одной жизненной форме. Последнее определяется тем, что к моменту возникновения простых группировок условия существования уже вызвали отмирание многих видов, для которых эти условия оказались неблагоприятными, а с другой стороны, еще не наступило время «сживания» друг с другом растений, относящихся к разным жизненным формам. Простые группировки обычно образованы некоторыми из видов, входивших в состав пионерных группировок. ПРИМЕР. На днищах высохших озер это часто марь (Chenopodiuт chenopodioides), астра солончаковая (Aster tripolium), в чистых простых группировках солерос (Salicornia herbacea), шведка (Suaeda maritiтa), петросимония трехтычинковая (Petrosimonia triandra), бассия иссополистная (Bassia hyssopifolia) в чистых и смешанных простых группировках. Простые смешанные группировки , существующие весьма длительное время – сообщества однотипных (например, накипных) лишайников на камнях. Простыми группировками обычно представлена бурьянистая стадия залежи. Сложная группировка – следующая за простой группировкой стадия развития фитоценоза. Она характеризуется следующими признаками: – видовой состав не вполне постоянен, – сообщество незамкнутое – новые виды могут легко в него проникать; – виды распространены еще не диффузно, хотя в скопления особей одного вида могут проникать особи других видов; – намечаются ярусы; – взаимное влияние растений становится еще более заметным; – обычно образована несколькими видами различных жизненным форм. ПРИМЕР. Зарастающая долина в районе строительства Уссурийской ТЭЦ. Сложную группировки здесь образуют рогоз (в мочажинах), донник, осоки разных размеров, мелкотравье. Растительный покров разрежен, но уже наметились ярусы: – рогоз – до 1 м, донник, лебеда, полынь, ослинник – 0,7–0,8 м, осоки – 0,4–0,5 м, мелкие злаки и травы высотой не более 10 см. Стадия замкнутого фитоценоза– следующая стадия развития фитоценоза характеризуется: – крайне затрудненным проникновением в него новых видов. – равномерное, не слишком густое распределение особей отдельных видов. – групповой рост – исключение. Представлена двумя формами сочетаний растений – фитоценозами зарослей и 2 и более ярусных фитоценозов. Заросли развиваются в условиях, в которых не может существовать сообщество из большого числа видов: сильная засоленность, сильная сухость, переувлажнение, высокая конкуренция, и т. п. Одноярусны. Ярус образован либо одним видом (чистые заросли), либо несколькими видами (смешанные заросли). ПРИМЕР. Травяные заросли (чистые): сообщество тростника (Phragтites coтmunis) без участия более низкорослых растении, сообщество пырея ползучего (Agropyron repens), и др. Пример травяных смешанных зарослей – сообщество камыша Табернемонтана (Scirpus tabernaeтontani) и рогоза узколистного (Typha angustifolia), заросли крапивы и лебеды на заброшенных площадках из–под удобрений. Кустарниковые заросли – саксаульники в пустыне Сахара(?), кедрово– и ольховниково–стланиковые – на Крайнем Северо–Востоке, ивняков и облепиховые в Южном Приморье, чозениевые в среднем возрасте на Дальнем Востоке. Сфагновые болота тоже можно считать моховыми зарослями – сфагнума. Многоярусный фитоценоз (простой из 2–х ярусов, сложный – из более, чем 2–х ярусов), развивающийся не в столь суровых условиях, как заросли. Это все типы лугов (пойменные, суходольные, залежные), все лесные сообщеста. Не во всех случаях фитоценоз обязательно проходит последовательно все перечисленные стадии: – пионерная группировка > простая группировка > сложная группировка > заросли или сложный фитоценоз. Этот путь бывает и проще, и сложнее. ПРИМЕР. На скалах: нередко пионерная группировка сине–зеленых водорослей >пионерная группировка лишайников > смешанная или простая группировка лишайников > смешанная длительно существующя заросль лишайников > сложный фитоценоз с участием лишайников, мхов > сложный фитоценоз цветковых растений. На освободившемся из–под воды днище пресного озера: смешанная пионерная (гигро) группировка > чистая пионерная группировка (ксерофит) > чистая простая группировка > смешанная простая группировка > сложной группировка и сложный фитоценоз. В других случаях чистую группировку сменяет чистая заросль, которая существует на этом участке неопределенно долгое время. Таким образом, путь развития фитоценоза разнообразен: более длинный и более короткий, включает то одни, то другие стадии. Но во всех случаях развитие его идет от раздельно–группового сложения к диффузному, от несомкнутого покрова к сомкнутому, от незамкнутого покрова к замкнутому. г) Стадии в развитии растительного покрова по В.Н. Сукачеву В.Н. Сукачев (1938, 1964 и др.) выделял следующие стадии формирования фитоценоза: 1. Отсутствие фитоценоза (соответствует пионерной группировке в начальной стадии ее существования). 2. Открытый фитоценоз (соответствует пионерной группировке в значительной части периода ее существования и простой группировке). 3. Закрытый невыработавшийся фитоценоз (соответствует сложной группировке). 4. Выработавшийся фитоценоз. Из других авторов изучением начальных стадий развития фитоценоза занимались Л. Г. Раменский (1938), А. А. Шахов (1941, 1946), Д. Уивер и Ф. Клементс (Weaver апd Clements, 1938). Методология изучения сукцессий Важно выяснить, как пойдет развитие растительного покрова на разных субстратах и при разном климате в тех случаях, когда человек вносит существенные изменения в состав растительного покрова, тем самым оказываясь причиной прохождения фитоценозом определенных стадий развития. Для решения последней задачи необходим эксперимент на постоянных пробных площадях. Стадии развития фитоценоза можно исследовать двумя путями. 1. Подбор возрастных рядов. Сравнивают участки растительности, расположенные рядом и находящиеся на разных стадиях развития. Детально описывая эти участки и делая зарисовки расположения отдельных экземпляров растений, получают представление и о сменах растительности во времени. 2. Наземный мониторинг. Можно проводить наблюдения на площадках, существующих длительное время (постоянные квадраты). На этих площадках, закладывающихся на участках, растительность которых представляет определенные стадии развития фитоценозов, подсчитывают количество или даже зарисовывают проекции отдельных экземпляров растений. Подсчеты и зарисовки повторяют в определенные сроки. Они помогают выяснить картину постепенного изменения растительного покрова через месяц, через год, через 5, 10 лет и т. д. Движущей силой автотрофных сукцессий является солнечная энергия, усваиваемая растениями–продуцентами и передаваемая по пищевым цепям консументам и редуцентам. Однако подобно тому, как существуют гетеротрофные экосистемы, возможны и гетеротрофные сукцессии (их называют также деградационными). Эти сукцессии происходят при разложении мертвого органического вещества (детрита): трупа животного, «лепешки» экскрементов коровы, упавшего ствола дерева, лесной подстилки и т.д. В гетеротрофных сукцессиях происходит «эстафета» биоты, которая представлена беспозвоночными, грибами и бактериями. Гетеротрофная сукцессия в опавшей хвое сосны продолжается около 10 лет (Бигон и др., 1989). Поскольку опавшая хвоя постоянно покрывается новыми слоями опада, то изучение лесной подстилки от ее верхней границы до почвы позволяет судить об изменении биоты во времени. Сукцессия происходит постепенно, тем не менее ее можно условно разделить на три стадии: – первая. Длится около 6 месяцев, в течение которых происходит первый этап разложения хвои. Впрочем, до 50% живых сосновых хвоинок уже поражено грибом Coniosporium, который открывает эту сукцессию. После опадения хвои этот гриб быстро исчезает, и на ней поселяются Fusicoccum и Pullularia. В конце стадии массово развивается Desmazierella; – вторая. Длится два года. В число участников сукцессии, кроме Desmazierella, включаются Sympodiella и Helicoma, к которым добавляются почвенные клещи; – третья. Наиболее продолжительная, которая длится 7 лет. Основными деструкторами хвоинок становятся почвенные животные – ногохвостки, клещи и олигохеты–энхитреиды. Хвоя спрессовывается, после чего интенсивность разложения резко снижается и сукцессия вступает в стадию «климакса». Другой пример – сукцессия состава насекомых–ксилофагов, участвующих в разложении древесины. Различаются пять стадий этой сукцессии (Кашкаров, 1944) со своим населением детритофагов: живой древесины, ослабленной древесины, мертвого целого дерева, частично разложившейся древесины, полностью разложившейся древесины. Гетеротрофную сукцессию можно продемонстрировать в эксперименте на сенном растворе, где вначале расцветает пышная культура разнообразных бактерий, которые при добавлении прудовой воды сменяются простейшими из родов Hypotricha, Amoeba, Vorticella. После того, как ресурсы исчерпываются, сукцессия останавливается, а участвовавшие в ней организмы переходят в покоящееся состояние. Вторичные сукцессии представляют собой восстановительные смены. Они начинаются в том случае, если в уже сложившихся сообществах частично нарушены установившиеся взаимосвязи организмов, например удалена растительность одного или нескольких ярусов (в результате вырубки, пожара, вспашки и т. д.). Смены, ведущие к восстановлению прежнего состава ценоза, получили в геоботанике название демутационных. Примером может служить демутация залежей в Абаканских степях, происходящая в четыре основных этапа: 1) преобладание однолетних сорняков – на 1–2–й год после вспашки; 2) господство «крупного бурьяна», преимущественно разных полыней – на 3–4–й год; 3) вытеснение их корневищными злаками (пыреем и др.) и появление большого количества бобовых – начиная с 5–го года; 4) установление господства степных дерновинных злаков с появлением ковыля – на 11–12–й год после вспашки. Восстановительные смены совершаются быстрее и легче, чем экогенетические, так как в нарушенном местообитании сохраняются почвенный профиль, семена, зачатки и часть прежнего населения и прежних связей. Демутации не являются повторением какого–либо этапа первичных сукцессии. Суть понятий сингенез, эндоэкогенез и гологенез а) На самых первых этапах развития сообщества преобладал процесс, который В.Н. Сукачев (1942) назвал сингенезом. Это процесс первоначального формирования растительного покрова, связанный с вселением растений на данную территорию, их приживанием (эцезисом), а затем и конкуренцией между ними из–за средств жизни. Затем начинается другой процесс, названный В. Н. Сукачевым эндоэкогенезом. Это процесс изменения фитоценоза под влиянием среды, изменененной им самим. Эндоэкогенез постепенно усиливается и, в конце концов, становится основным процессом, определяющим ход смен фитоценоза. На эти два процесса налагается третий, названный В. Н. Сукачевым (1954) гологенезом. Это «процесс изменения растительного покрова под влиянием всей географической среды или отдельных ее частей: атмосферы, литосферы и т.п., т.е. изменения более крупного единства, в состав которого входит данный биогеоценоз. Все три процесса идут одновременно, но на разных стадиях развития преобладающее значение приобретает один из них. Несомненно, сингенез господствует только на начальных стадиях развития фитоценоза, а затем главенствующая роль переходит к эндоэкогенезу. Гологенетический процесс протекает постоянно, но, очевидно, в переломные моменты геологической истории Земли роль его усиливается. Такой ход развития фитоценоза продолжается большее или меньшее время до тех пор, пока какие–то внешние силы, случайные по отношению к ходу развития фитоценоза, резко не нарушают его. Тогда смена, вызванная внутренним развитием самого фитоценоза (эндодинамическая), прерывается, и начинается смена, вызванная внешним толчком (экзодинамическая). Исходя из сказанного выделяются два основных типа смен фитоценозов (Сукачев, 1928): 1. эндодинамические, происходящие в результате постепенного развития самого фитоценоза, меняющего среду и при этом изменяющегося; основную роль играют внутренние особенности сообщества. 2. экзодинамические (Сукачев, 1928; Лавренко, 1940), или стихийные (Ярошенко, 1953), или внезапные (Ярошенко, 1961), возникающие под непредвиденным воздействием внешних факторов. Причины возникновения сукцессий (смен) растительного покрова весьма разнообразны. ПРИМЕРЫ эндодинамических смен – процессы заболачивания лесов в результате изменений в их напочвенном покрове, процессы зарастания водоемов, развития растительного покрова на скалах и обсохших днищах водоемов и т. д. Экзодинамические смены – смены растительного покрова на вырубках, залежах, пожарищах, участках, засыпанных лавиной или залитых селевым потоком, и т.д. б) Классификация типов сукцессий по П.Д. Ярошенко П. Д. Ярошенко (1961), разрабатывая теорию сукцессий, составил более подробную следующую классификацию типов смен: А. Природные смены. Последовательные: а) эндоэкогенетические (включают и сингенетические смены В.Н. Сукачева), б) гологенетические. Внезапные смены: а) климатогенные, б) эдафогенные, в) биогенные. Б. Антропогенные смены: а) последовательные, б) внезапные. ПРИМЕРЫ разных смен. Климатогенные смены – смены типчаково–серополынных фитоценозов серополынно–типчаковыми при изменении климата в сторону большей влажности; эдафогенные смены –– заселение растительностью вновь возникших в результате естественных причин водоемов, зарастание вновь возникших под влиянием эрозионной и аккумулятивной деятельности обнаженных или полуобнаженных участков ("лунных ландшафтов") и т.д.; зоогенные смены – результат подгрызающей и роющей деятельности животных, а также подпруживания рек бобрами; пирогенные смены – изменения растительного покрова в результате пожаров; антропогенные смены – следствие вырубки лесов, распашки территории, сенокошения, 'выпаса домашних животных и т. п.). Антропогенные смены нецелесообразно отделять от природных. Во–первых, нередко действие человека и действие природного фактора аналогичны В данной системе природные внезапные смены и антропогенные смены соответствуют экзодинамическим сменам В.Н. Сукачева, поскольку с точки зрения существования растительного сообщества безразлично, какими обстоятельствами вызвана серия смен: вырубкой леса, его уничтожением в результате пожара или в результате действия насекомого–вредителя, хотя ход процессов при этом будет неодинаков. Любая смена, начавшаяся под влиянием внешних причин (экзодинамически), в дальнейшем постепенно переходит в эндодинамическую, так как ее ход во все большей степени определяется воздействием фитоценоза на среду. Если вновь не повторятся воздействия экзогенных факторов на фитоценоз. в) Кратковременные, вековые и филоценогенетические смены фитоценозов По длительности сукцессии подразделяются на кратковременные (частные), вековые (общие) и филоценогенетические (эволюция фитоценозов). Все перечисленные типы смен очень тесно сплетаются друг с другом и накладываются один на другой. В группу кратковременных, или частных, смен объединяют эндодинамические и экзодинамические. Им противопоставляются вековые (Лавренко), или общие (Ярошенко), смены. Последние протекают значительно медленнее и охватывают территорию, занимаемую не одним или немногими связанными друг с другом фитоценозами, а обширные территории. Между кратковременными и вековыми сменами существуют переходы, хотя в общей форме можно сказать, что в результате кратковременных смен вырабатываются растительные ассоциации и формации в пределах географических зон, а в результате вековых смен растительность одной зоны сменяется растительностью другой. ПРИМЕРЫ кратковременных смен: смены сосняков из сосны могильной – сухими дубняками, хвойно–широколиственных лесов – смешанными широколиственными лесами; вековых смен: изменения растительного покрова на севере европейской части СССР после отступления ледника и на юге Дальнего Востока – после отступления–наступления моря. Наконец, различают филоценогенез, или эволюцию фитоценозов. г) Эволюция фитоценозов. Примеры филоценогенетических сукцессий Эволюция фитоценозов, как указывал В. Н. Сукачев (1954), слагается из двух процессов: выработки структуры ценоза (филоценогенез) и видообразования, идущего в связи с эволюцией ценозов (флорогенез). Эволюции растительных сообществ совершается также путем естественного отбора, который В. В. Ревердатто (1935) предложил называть естественным отбором второго порядка (в отличие от естественного отбора первого порядка – отбора особей в процессе видообразования. Благодаря естественному отбору существует определенное число фитоценозов, а не бесчисленное количество комбинаций, сочетаний растений. В процессе эволюции выработались как растительные ассоциации, так и более крупные единицы растительного покрова вплоть до типов растительности. В качестве примеров можно привести примеры частных смен фитоценозов, происходящих в различных условиях. Существует два способа зарастания водоемов: со дна и с поверхности. Детали этого процесса в разных районах и в водоемах разных типов неодинаковы, но общий ход его для каждого из этих способов один и тот же. О ходе сукцессий в водоемах можно судить, исследуя отложения сообществ, сменявших друг друга на данной территории, или путем изучения поясов растительности, сменяющих друг друга по мере изменения условий существования в пространстве. Процесс зарастания водоема со дна При зарастании пресноводных стоячих водоемов сo дна, чаще всего при пологих берегах, наблюдается несколько стадий. Связь между стадиями заключается в оседании на дно остатков организмов и повышения уровня дна. Это делает возможным продвижение пояса больших растений – макрофитов, связанных с определенными глубинами, от первоначального берега к центру водоема и приводит к смене озера болотом. На болоте также по мере накопления торфа и изменения условий существования одна растительная ассоциация сменяет другую. ПРИМЕР № 1. При зарастании водоема со дна обычно наблюдаются следующие стадии (Лепилова, 1934): 1. Господство планктона. Глубины еще слишком значительны, чтобы зеленые растения–макрофиты могли здесь обитать. 2. Подводные луга с глубины 5, иногда 6 м, до нижнего предела существования растительности макрофитов. Здесь растут водяные мхи скорпидиум (Scorpidiuт), каллиергон (Calliergon) и др., виды харовых водорослей (Chara), иногда сюда спускается элодея канадская (Elodea canadensis). 3. Заросли широколиственных рдестов, развитые на глубинах 3–5 м (иногда до 6 м). Образованы погруженными в воду растениями, выносящими на поверхность воды или поднимающими над ее поверхностью свои цветки для опыления. Здесь наиболее часты рдесты пронзеннолистный и блестящий (Potaтogeton perfoliatus, P. lucens) и др. 4. Заросли растений с плавающими листьями, развитые на глубинах 2–3 м, иногда более (до 5 м), обычно в защищенных от ветра местах: в заливах, заводях, старицах и т. д. Для этих зарослей характерны кубышки (Nuphar), кувшинки (Nyтphaea), рдест плавающий (Potaтogeton nataпs). 5. Пояс камыша и тростника. Развит с глубины 0,5 до 2–3 м, редко глубже. Здесь растут виды, поднимающие свои стебли высоко над поверхностью воды. Это камыш озерный (Scirpus lacustris), тростник обыкновенный (Phragтites coттuпis), рогозы (Typha), манник водяной (Glyceria aquatica), тростянка (Scolochloa) и др. На этой стадии дно водоема покрыто мощным слоем ила. 6. Прибрежная растительность мелководий. Обитает от уреза воды до глубины 0,5 м, т.е. до пояса камыша и тростника. Характер дна на этой стадии очень изменчив. Оно бывает илистым, песчаным и каменистым (песчаное и каменистое дно обычны в северных, слабо зарастающих озерах). Характерны для этой стадии осоки, главным образом осока вздутая (Саrех rostrata), рдест разнолистный (Potaтogetoп heterophyllus), хвощ приречный (Equisetuт heleocharis). 7. Прибрежная растительность. Занимает участки, расположенные выше уреза воды, но по временам заливаемые водой при высоком ее уровне или при сильном прибое. Дно здесь может быть илистым, торфянистым, песчаным, каменистым, глинистым и т. д. Растения, характеризующие эту стадию, относятся к числу земноводных, иногда болотных. Здесь характерны вех (Cicuta), ситняги болотный и игольчатый (Heleocharis palustris, Н. acicularis), лютик стелющийся (Raпuпculus reptaпs), осоки (Саrех) и др. Дальнейшее развитие сукцессий может привести к возникновению сообществ кустарников и деревьев. Частными вариантами сукцессии являются смены, которые по продолжительности точно совпадают с каким–либо сезоном, а на следующий год начинаются сначала. Эти «сезонные» сукцессии выглядят как обычная фенологическая смена форм, но природа их иная. Например, в планктоне северных морей весной массовое размножение водорослей сменяется волной размножения рачков–калянусов, затем в большом количестве появляются гребневики, а после них – некоторые виды рыб. Эти смены происходят как постепенное развертывание в течение короткого северного лета единой гигантской трофической цепи организмов, так как каждая последующая группа питается за счет предыдущей. Если же в силу неблагоприятной ситуации не возникает, например, весной массовое размножение фитопланктона, то и все последующие группы будут угнетены, несмотря на благоприятные сочетания любых прочих факторов в период обычного появления группы.
ПРИМЕР № 2. Зарастание со дна заводей в Полабье (Чехословакия): 1) водное сообщество Myriophylleto–Nupharetum; 2) сообщество Parvocharetum, подготовливающее заполнение заводи подстилкой, пропитанной известняком; 3) заросли тростника – Phragтitetum; 4) заросли Cladiuт–Cladietum marisci; 5) пояс дерновинных осок (сообщество из группы Magno–caricetum); 6) заросли Schoeпus–Schoenetum nigricantis; уровень грунтовых вод находится ближе к поверхности, как и в 7) сообществе Moliпia–Molinietum caficeto–sumpaniceae, которое вытесняется на крепкой почве стадией 8) верб и крушины Franguleto–Salicetum, подготовляющей почву для 9) ольшаника – ассоциации Alпus glutiпosa–Dryopteris thelypteris.
Процесс зарастания водоемов с поверхности ПРИМЕР № 3. Совершенно иначе зарастают водоемы с поверхности (заболачивание путем нарастания) При этом с берега на поверхность воды наплывает зыбучий ковер из мхов и некоторых цветковых растений, с мощными корневищами (рис. 2). Такой ковер называется сплавиной, или зыбуном. Сплавина образуется только в защищенных от ветра местах с относительно круто уходящим в глубь от берега дном. Сплавина разрастается, от ее нижней поверхности отрываются куски торфа, перегнившие и полуперегнившие, остатки растений и на дне образуется мощный слой полужидкого ила. Постепенно весь водоем заполняется кашицеобразной массой, а с поверхности его покрывает сплошной слой сплавины, в котором просвечивают «окна» – отверстия или участки, где сплавина очень тонка. Попав в такое «окно», человек или животное погибает в трясине. После заполнения водоема илом на его месте развивается болото.
Процесс образования болот В зависимости от условий местности дальнейшее развитие заросших водоемов приводит или к возникновению лесной растительности, или к образованию верхового болота, лишенного связи с грунтовыми водами, имеющего атмосферное питание и характеризующегося господством сфагновых мхов . Причины заболачивания суши различны. 1. Материковые луга могут заболачиваться в результате уплотнения почвы под влиянием весеннего выпаса скота. 2. Существенная причина заболачивания материковых лугов – обеднение почв. Питательные вещества выносятся животными с поедаемыми растениями или вывозятся со скошенным сеном. Более требовательные к питательным веществам ценные травы постепенно исчезают, появляются растения, мирящиеся с бедностью почвы, среди которых большую роль играют мхи.
3. После вырубки леса или после лесного пожара в районах с неглубоким уровнем грунтовых вод, также начинаются процессы заболачивания, которые связаны с подъемом грунтовых вод по почвенным капиллярам, вследствие прекращения вытягивания воды из почвы деревьями. 4. На севере лесной зоны заболачивание может вызвать естественная смена зеле–ных гипновых мхов кукушкиным льном и сфагнумом. Кукушкин лен удер–живает большое количество воды, что влияет на степень увлажнения почвы. Когда влажность становится чрезмерной для кукушкина льна, он сменяется сфагновыми мхами, а те, способствуя сохранению вечной мерзлоты, еще более способствуют заболачиванию. Сфагновые мхи появляются сначала небольшими пятнами, а затем постепенно завоевывают господство, могут и не предваряться зелеными мхами.
Зарастание горных территорий с каменистыми субстратами Ход и характер сукцессий определяется составом пород, образующих скалы, крутизной и экспозицией скалистого склона. Очень медленно происходит завоевание растительностью скал. На обнаженных скалах сначала поселяются бактерии, синезеленые и зеленые водоросли, затем лишайники. Лишь спустя очень долгое время, когда в результате выветривания и деятельности бактерий и лишайников на поверхности скалы местами образуется незначительный почвенный слой, здесь могут развиваться мхи. После поселения мхов накопление частиц почвы происходит быстрее. Спустя еще некоторое время на скалах появляются папоротникообразные и цветковые растения, сначала по трещинам и расщелинам, где скапливаются органические вещества и вода, а затем на их поверхности.
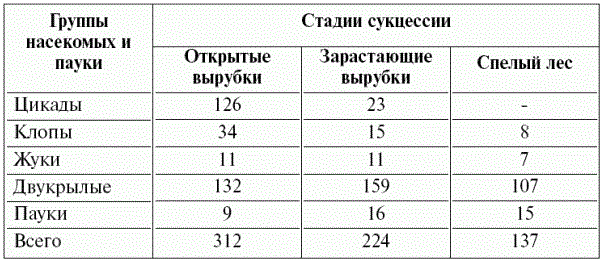
ПРИМЕР № 4. На известняковых скалах в районе Праги, на склонах южной экспозиции, сменяют друг друга следующие сообщества: 1 – папоротник Aspleпiuт rutaтuraria в трещинах скал; 2 – сообщество Festuca duriuscula–Seseliesseum на сильно скелетной неглубокой почве, которую оно скрепляет, обогащает гумусом и подготовляет для следующего сообщества; 3 – сообщество Carex huтilis–Festuca sulcata, которое преобладает на более пологих ступенчатых склонах с почвой менее скелетной и более мощной; 4 – на более глубоких почвах пологих склонов развито замкнутое сообщество Eestuca vallesiaca–Erysiтuт crepidifoliuт, в которое проникают 5 – кустарники, особенно степная вишня – Prunus fruticosa, предшествующая 6 – грабовому лесу с дубом краснеющим или теплолюбивой дубраве (Quercetum–Carpinetum primuletosum). На верхушках скал стадии 4–6 мозаично сменяют друг друга, внешнеи напоминая (образуя) «лесостепь». Местные причины могут изменить ход сукцессии или привести к фрагментарному выражению ее стадий (2–6). На склонах северных и западных экспозиций сукцессии происходят иначе, чем на южных. Восстановительные сукцессии по своему характеру мало отличаются от первичных, но, как отмечалось, протекают в экосистемах, которые частично или полностью нарушены внешним воздействием (как правило, деятельностью человека). Они обычно протекают быстрее, чем первичные, на их скорость влияет степень сохранности экосистемы и наличие источников диаспор для ее восстановления. Классический пример такой сукцессии – восстановление степи или леса на месте заброшенной пашни. Примерно 150 лет назад основными системами земледелия в России были залежно–переложная и подсечно–огневая (соответственно в степной и лесной зонах). Участок земли использовался как пашня 5–10 лет, после чего забрасывался, т. к. почва истощалась и обильно развивались сорняки, представлявшие первую стадию восстановительной сукцессии уже под пологом культурного растения. Контролировать сорняки при отсутствии тракторов и пестицидов человек не умел. Постепенно на заброшенном поле, через стадии полевых (сегетальных) сорняков, которые доминировали в первый год, и рудеральных видов, разраставшихся в последующие 3–5 лет, формировался степной травостой или вырастал лес. В ходе этой сукцессии восстанавливалось плодородие почвы, а сорные растения вытеснялись более мощными рудеральными, луговыми и лесными видами. Соответственно обогащалась и фауна. Восстановление растительности на залежах происходило достаточно долго – не менее 25 лет. Человек научился ускорять этот процесс. Дж. Кертис (J. Curtis) в двадцатых годах прошлого столетия значительно быстрее восстанавливал прерии за счет «искусственного семенного дождя» – смеси семян, собранных на сохранившихся участках прерии. Восстановление лугов высевом смеси семян, собранных в естественных луговых сообществах, практикуется сегодня в Англии. Ставропольский ботаник Д. Дзыбов разработал экономичный способ ускорения восстановительной сукцессии путем рассева сена с целинного степного участка на вспаханную почву. Семена высыпаются в почву, и сукцессия восстановления степи резко ускоряется: к пятому году в такой «агростепи» есть уже до 80% видов растений целинной степи. Для ускорения восстановительных сукцессий экосистем тундры на Аляске, нарушенных при добыче нефти, применяли азотные удобрения. Восстановительные сукцессии активно протекают не только на залежах, но и в посевах многолетних трав. Это позволяет использовать старовозрастные посевы многолетних трав для повышения биологического разнообразия сельскохозяйственных экосистем. Само собой разумеется, что в ходе восстановительных сукцессий меняется вся гетеротрофная биота экосистемы. В литературе приводятся данные об изменениях фауны птиц, грызунов, насекомых. Сукцессия состава населения птиц изучалась в прериях США (Одум, 1986). Количество видов гнездящихся птиц менялось от 15 до 239, причем на разных стадиях сукцессии состав птичьего населения существенно менялся: – на первой стадии (первые три года), когда доминировали травянистые растения, число видов птиц менялось от 15 до 40 видов, причем доминировали саванный воробей и луговой трупиал; – на второй стадии – кустарников, которая продолжалась 22 года, орнитофауна возросла до 136 видов, причем наиболее массовыми были: американская славка, овсянка, желтогрудая славка; – на третьей стадии – соснового леса, которая представляла 35–100 годы сукцессии, орнитофауна была самой богатой и достигала 239 видов. Самыми массовыми были древесница, тонагра, тиранн, верион желтолобый; – на заключительной стадии – дубово–гикориевого леса, которая формируется через 150–200 лет после забрасывания пашни, разнообразие птичьего население снизилось до 228 видов. К видам соснового леса добавляются американская кукушка, еще два вида древесницы и тиранн зеленый. Аналогичные данные о динамике фауны при восстановлении леса получила М.Н. Керзина (1956). Так восстановление ельника (Костромская область) сопровождалось изменением фауны грызунов и насекомых. На стадии открытой лесосеки (1–2 года после вырубки) фауна грызунов была представлена видами из рода Microtus, на смену которым при восстановлении леса приходили типичные лесные виды грызунов из рода Clethrionomys, причем на средней стадии сукцессии эти виды сочетались. Сходный характер имела и динамика насекомых (табл. 16). В целом энтомофауна обеднялась за счет резкого уменьшения числа цикад, уменьшалось количество особей других групп, исключая пауков, количество которых увеличивалось.
Таблица 16 – Динамика численности основных групп насекомых при восстановлении еловых, елово–пихтовых и сосновых лесов (на 100 взмахов сачком; по М.Н. Керзиной, 1956)
Распространенным вариантом вторичной восстановительной сукцессии является постпастбищная демутация. При снижении пастбищной нагрузки начинается процесс восстановления пострадавших от выпаса высоких трав: овсяницы луговой, ежи сборной и костреца безостого – на лугах и ковылей – в степях. Патиенты–пастбищники (подорожники, одуванчик, лапчатка гусиная, клевер ползучий на лугу; полынь австрийская и типчак в степи) при отсутствии сильного выпаса теряют свои конкурентные преимущества и резко снижают обилие. Аллогенные сукцессии вызываются факторами, внешними по отношению к экосистемам. Такие сукцессии чаще всего протекают в результате влияния человека, хотя возможны и природные аллогенные изменения. Их пример – изменение состава экосистемы поймы в результате меандрирования реки и углубления ею базиса эрозии русла. В итоге уровень поймы повышается, а длительность заливания и количество наилка уменьшаются. В результате этого в экосистемах пойм умеренной полосы последовательно сменяют друг друга сообщества ивняков, тополевников, вязовых и липово–дубовых лесов и полностью меняется состав травянистых видов. Меняется и состав гетеротрофной биоты, так как растительные сообщества предоставляют им соответствующую кормовую базу. Кроме того, состав растительного сообщества отражает длительность затопления в период паводка, что во многом предопределяет возможность выживания разных видов насекомых, птиц, почвенной фауны и т.д. Наиболее распространенным примером аллогенной сукцессии является изменение экосистем злаковников (лугов и степей) под влиянием выпаса. При высоких пастбищных нагрузках снижается видовое богатство, биологическая продукция, биомасса и происходят изменения состава растительного сообщества и сопровождающей его фауны: на смену высоким и хорошо поедаемым растениям приходят низкорослые и плохо поедаемые (последние могут быть и высокорослыми, как, например, виды чертополоха – род Carduus). В степных экосистемах различаются стадии пастбищной дигрессии: ковыльная, типчаковая (с Festuca valesiaca или F. pseudovina), полынковая с господством Artemisia austriaca. На заключительных стадиях такой сукцессии происходит рудерализация и массово развиваются однолетники, которые используют для быстрого роста перерывы между циклами стравливания и условия ослабленной конкуренции с многолетниками, которые угнетены выпасом. Сегодня чрезвычайно распространенным и нежелательным процессом изменения водных экосистем является их эвтрофикация – изменение в результате поступления большого количества элементов минерального питания, в первую очередь фосфора. Основной причиной эвтрофикации является смыв удобрений с полей, а также стоки животноводческих ферм. В ходе сукцессии первыми гибнут диатомовые водоросли, вслед за ними – зеленые водоросли, которые вытесняются цианобактериями. Некоторые штаммы цианобактерий выделяют в воду токсичные вещества, которые вызывают гибель многих организмов. При опускании на дно они разлагаются редуцентами, что требует большого количества кислорода. В итоге в таком обедненном кислородом водоеме гибнет большинство видов рыб и макрофитов (в первую очередь таких требовательных к чистой воде, как сальвиния, водокрас лягушечий, горец земноводный). В то же время, роголистник, рогоз широколистный и ряски могут выдерживать достаточно высокий уровень загрязнения и сохраняться в такой эвтрофицированной экосистеме. Вокруг эвтрофицированного водоема ощущается дурной запах, в мелководье скапливается бурая пена, содержащая погибший планктон. Если количество стоков ограничено или они уже прекращены, водная экосистема сама может справиться с загрязнением – произойдет процесс деэвтрофикации, описанный в предыдущем разделе. Успешно противостоять эвтрофикации могут макрофиты, активно усваивающие элементы питания. Однако самоочистительная способность водных экосистем ограничена, и потому если стоки поступают длительное время и в большом количестве – они гибнут. От эвтрофицикации следует отличать отравление водных экосистем промышленными и бытовыми стоками, которые содержат токсичные вещества, например тяжелые металлы. Если поступление токсикантов ограничено, то экосистема может справиться и с ними: ядовитые вещества попадут в организмы ее обитателей, а после их смерти будут захоронены на дне. На дне водоемов Куйбышевского, Волгоградского и других водохранилищ накопился многометровый слой токсичных осадков, образующихся в процессе самоочищения. Однако, если поступит значительное количество токсичных веществ и тем более если они будут поступать регулярно, водная экосистема восстановиться не сможет. Другим примером аллогенной сукцессии является изменение состава экосистем под влиянием радиации. Они были изучены Р. Уиттекером и Г. Вудвелом (Whittaker, Woodwell, 1972) на радиационном полигоне о. Лонг (США). При повышении дозы радиации (использовался источник гамма–излучения) происходила сукцессия, которая была как бы зеркальным отражением сукцессии зарастания скал, описанной Ф. Клементсом: вначале гибли деревья, потом кустарники, травы, мхи, и при самых высоких дозах радиации сохранялись только почвенные водоросли. В районе Чернобыля после аварии сукцессия прошла первую стадию: в лесах, расположенных вблизи АЭС, усох древостой (однако спустя несколько лет он начал интенсивно восстанавливаться). Как правило, аллогенные сукцессии сопровождаются снижением продуктивности и биоразнообразия, хотя на первых стадиях сукцессии эти параметры могут возрастать. Травяные сообщества при умеренном выпасе, леса при некотором влиянии отдыхающих или водные экосистемы при легкой эвтрофикации имеют более богатый видовой состав, чем те же сообщества, не испытывающие внешних влияний. В некоторых случаях при аллогенной сукцессии возрастает продукция, но снижается видовое богатство. Это наблюдается при изменении лугов под влиянием минеральных удобрений: число видов в сообществах уменьшается в 2–2,5 раза. Причина тому – обострение конкуренции при повышении уровня обеспечения ресурсами. Так большой ущерб видовому составу европейских горных лугов на бедных почвах принесли мероприятия по их улучшению путем внесения минеральных удобрений. Подобным образом снижение видового богатства может сопровождаться возрастанием биологической продукции и при эвтрофикации водоемов.
Смены биогеоценозов могут быть прогрессивными и регрессивными. Под первыми обычно понимают такие смены, которые связаны с увеличением продуктивности и усложнением строения фитоценоза. Смены, связанные с уменьшением запасов растительной массы и упрощением строения фитоценоза, следует считать регрессивными. Так, заселение растительностью скал служит примером прогрессивной смены, а заболачивание лесов – пример регрессивной смены. Человек заинтересован в том, чтобы направлять смены растительного покрова в сторону прогресса. Знание направления, особенностей хода и темпа сукцессий не обходимо для управления этими процессами. Вопросы возобновления леса, развития растительности на пастбищах, озеленения терриконов и выемок и многие другие могут быть разрешены только после тщательного изучения сукцессий.
Своеобразный вариант смен представляютпоточные, или конвейерные, сукцессии. Они развиваются в подвижной среде: реках, потоках, круговоротах водных масс в океанах. Планктонные сообщества, увлекаемые водой, на разных стадиях развития оказываются в разных географических точках, а в одном и том же регионе постоянны по составу. Такой же конвейерный тип преобразований наблюдается, например, в лесной подстилке. В разных ее подгоризонтах: верхнем рыхлом слое опада, среднем спрессованном «ферментативном» подгоризонте и нижнем, полностью переработанном муллевом слое – существует постоянная значительная разница в многочисленном микронаселении. Но в каждой конкретной порции опада, поступающего сверху, происходит сукцессионная смена микроорганизмов, простейших, нематод и мелких членистоногих, обеспечивающая все более глубокое разложение растительных остатков. Опад вместе со всем населением постепенно занимает все более глубокое положение, захораниваясь под новыми поступлениями, а общая стратификация подстилки выглядит неизменной.
|