Тема 5.2. Устойчивость природных экосистем
Процесс сукцессии, по Ф. Клементсу, состоит из нескольких этапов: 1) возникновения незанятого жизнью участка; 2) миграции на него различных организмов или их зачатков; 3) приживания их на данном участке; 4) конкуренции их между собой и вытеснения отдельных видов; 5) преобразования живыми организмами местообитания, постепенной стабилизации условий и отношений. В настоящее время практически вся доступная жизни поверхность суши занята различными сообществами, и поэтому возникновение свободных от живых существ участков имеет локальный характер. Это или места, освободившиеся в результате отодвигания ледников, отступания уреза воды в водоемах, обвалов, эрозии и т. п., или возникшие в результате деятельности человека, например выноса наверх больших масс глубинных пород при разработке полезных ископаемых. Занос спор, семян растений, проникновение животных на освободившийся участок имеют большей частью случайный характер и зависят от того, какие виды есть в окружающих биотопах. Из числа видов, попавших в новое местообитание, приживаются лишь те, экологическая валентность которых соответствует данному комплексу абиотических условий. Прижившиеся виды постепенно занимают весь новый биотоп, вступая в конкуренцию друг с другом. В результате происходит перестройка видового состава и количественных соотношений разных форм. Параллельно идет процесс преобразования самого местообитания под влиянием развивающегося сообщества. Процесс завершается формированием более или менее стабильной системы с уравновешенным типом биологического круговорота. Сукцессии любого масштаба и ранга характеризуются целым рядом общих закономерностей, многие из которых чрезвычайно важны для практической деятельности человека. В любой сукцессионной серии темпы происходящих изменений постепенно замедляются. Конечным итогом является формирование относительно устойчивой стадии – климаксового сообщества, или климакса . Начальные, пионерные группировки видов отличаются наибольшей динамичностью и неустойчивостью. Климаксовые же экосистемы способны к длительному самоподдерживанию в соответствующем диапазоне условий, так как приобретают такие черты организации биоценозов, которые позволяют поддерживать сбалансированный круговорот веществ. В ходе сукцессии постепенно нарастает видовое многообразие. Это ведет к усложнению связей внутри ценоза, разветвлению цепей питания и усложнению трофической сети, умножению симбиотических отношений, усилению регуляторных возможностей внутри системы. Тем самым уменьшается вероятность слишком сильного размножения отдельных видов и снижается степень доминирования наиболее массовых форм. Чрезвычайное увеличение численности отдельных видов возможно чаще всего на начальных этапах развития сукцессионных серий, когда еще в недостаточной мере сложилась система взаимного регулирования. В незрелых сообществах, т. е. находящихся в начале сукцессионных рядов, преобладают мелкоразмерные виды с короткими жизненными циклами и высоким потенциалом размножения, специализированные на быстром захвате освободившегося пространства. Они обычно обладают широкими расселительными возможностями, позволяющими им первыми проникать на незанятые участки, но малоспособны к конкуренции и длительному удержанию за собой пространства. Постепенно в развивающихся сообществах появляются и закрепляются более крупные формы с длительными и сложными циклами развития. Нарастание экологического разнообразия ведет к более четкому распределению групп организмов по экологическим нишам. В растительном покрове становится сильнее выражена ярусность и мозаичность, создающие основу пространственной структуры наземных экосистем. Усиливается зависимость успешного существования одних видов от биохимических выделений, роста или поведения других: преобладание конкурентных взаимоотношений сменяется преобладанием мутуалистических и трофических зависимостей. В результате сообщества приобретают известную степень автономности и независимости от окружающих условий, не подчиняя свою жизнь флюктуациям внешней среды, а вырабатывая собственные эндогенные ритмы. Не меньшие преобразования происходят и в энергетическом балансе системы (рис. 157). С энергетических позиций сукцессия – такое неустойчивое состояние сообщества, которое характеризуется неравенством двух показателей: общей продуктивности и энергетических трат всей системы на поддержание обмена веществ. 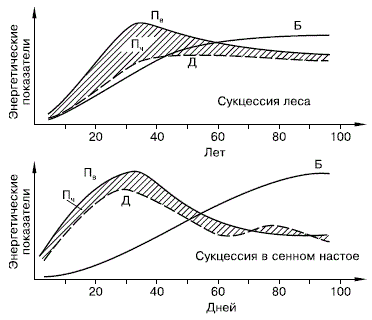
Рисунок 157 – Изменение энергетических показателей сообществ при развитии соснового леса (вверху) и лабораторной культуры простейших в сенном настое (внизу) (по Ю. Одуму, 1975):
Пв – продукция валовая; Д – траты на дыхание; Пч – продукция чистая (заштриховано); Б – биомасса
В ходе сукцессии общая биомасса сообщества сначала возрастает, но затем темпы этого прироста снижаются, и на стадии климакса биомасса системы стабилизируется. Это происходит потому, что на первых этапах сукцессии, когда видовой состав сообществ еще беден и пищевые цепи коротки, не вся часть прироста растительной массы потребляется гетеротрофами. Таким образом, относительно высока чистая продукция сообщества, которая идет на увеличение его биомассы. Накапливаются как общая масса живых организмов, так и запасы мертвого, неразложившегося органического вещества. В зрелых, устойчивых экосистемах практически весь годовой прирост растительности поступает и расходуется в цепях питания гетеротрофами, поэтому чистая продукция биоценоза, его «урожай», приближается к нулю. Биогенные элементы растения сначала черпают из запасов почвы, но постепенно, по мере истощения этих запасов и накопления в системе мертвого органического вещества, разложение его становится основным источником минерального питания растений, и круговороты биогенных веществ из открытых превращаются в закрытые. Неравенство трат энергии на образование первичной продукции и суммарный метаболизм (дыхание) сообщества проявляется в тех сукцессиях, в которых участвуют только гетеротрофные организмы. В этом частном случае первичная продукция равна нулю, и если приток мертвого органического вещества, за счет которого существует сообщество, не восполняет запасы в системе, то сообщество вскоре истощает свою энергетическую базу. С удлинением цепей питания увеличивается эффективность использования поступающей энергии, иначе – КПД всей системы, так как одна и та же порция энергии идет на поддержание большого количества биомассы. Знание этих закономерностей имеет большое значение в практической деятельности человека. Изымая избыток чистой продукции из биоценозов, находящихся в начале сукцессионных рядов, мы задерживаем сукцессию, но не подрываем основу существования сообщества. Вмешательство же в стабильные, климаксовые системы, с большой полнотой расходующие энергию на «свои» нужды, неминуемо вызывает нарушения сложившегося равновесия. Пока нарушения не превышают самовосстановительной способности ценоза, возникающие демутационные смены могут вернуть его к исходному состоянию. Этим пользуются, например, при рациональном планировании рубок леса. Но если сила и частота воздействия выходят за рамки этих возможностей, то первоначально устойчивое, богатое видами сообщество постепенно деградирует, сменяясь производными с малой способностью к самовозобновлению. Вырубка леса на локальных участках с оставлением части территории под коренными типами лесной растительности вызывает ускоренные сукцессии, исходные фитоценозы восстанавливаются за относительно короткий срок – немногие десятилетия. Сплошные же рубки на больших площадях, особенно если используется мощная корчевальная техника, полностью разрушают не только лесное сообщество, но и весь почвенный покров, эволюция которого шла тысячелетия. В этом случае сукцессии приводят к иным, упрощенным типам сообществ, и на месте лесов возникают пустоши, болота или другие малопродуктивные экосистемы. Таким образом, сообщество не может одновременно сочетать два противоположных свойства: быть высокостабильным и давать большой запас чистой продукции, который можно было бы изымать без вреда для самого ценоза. Сукцессии, протекающие при разложении органического вещества в почве, лежат в основе биологического круговорота. Это естественные регуляторные процессы, восстанавливающие нормальное состояние нарушенных почв. Такие проблемы века, как подрыв естественного плодородия в результате нарушения процессов образования гумуса, загрязнение среды ядохимикатами и органическими отходами, эрозия, «утомление почв» и другие отрицательные явления, возникли вследствие ослабления регуляторных возможностей почв. Стабильность экосистем, надежность протекания биологического круговорота веществ основаны на видовом разнообразии и полноте сукцессий. Первые переселенцы, которые приживаются на новом участке – это организмы толерантные к абиотическим условиям нового для них местообитания. Не встречая особого сопротивления среды, они чрезвычайно быстро размножаются (саранча, эфемерная растительность), то есть на ранних этапах эволюции экосистемы преобладает г– стратегия (рост численности), но постепенно возрастает видовое разнообразие за счет достаточно быстрой смены и увеличения количества популяций и начинает возрастать значение IC–фактора (ограничитель роста). Увеличение видового разнообразия приводит к усложнению связей внутри сообщества, умножению симбиотических связей, снижению чрезмерной рождаемости и доминирования массовых видов. Наконец действия г– и К–факторов уравновешиваются и сообщество развивающейся серии становится стабильным, или климаксным – это самоподдерживающееся сообщество, находящиеся в равновесии с физическим местообитанием. Развивающееся сообщество преобразует и само местообитание. На первых этапах для растительных форм первостепенное значение имеют почвенные биогенные элементы. Но черпать их из запасов почв до бесконечности невозможно и по мере истощения этих запасов разложение отмершей органики становится основным источником питания минеральными веществами биогеохимического круговорота. Однако такой круговорот возможен лишь в автотрофной системе, черпающей энергию от солнца. Другое дело – гетеротрофная сукцессия, когда приток мертвого органического вещества не восполняет запасы, то есть первичная продукция равна нулю, и участвуют в сукцессии только гетеротрофные организмы. В этом случае количество энергии не добавляется, а уменьшается, и система прекращает свое существование – все организмы погибают или, в лучшем случае, переходят в покоящиеся стадии (сукцессия в гниющих стволах деревьев, в трупах животных, фекалиях). Такая сукцессия должна ассоциироваться, по мнению Ю. Одума, с эксплуатацией залежей горючих полезных ископаемых человеком. На ранних стадиях сукцессионной серии чистой продукции получается значительно больше и при ее изымании человеком сукцессия только приостанавливается, но основа продуктивности на этих этапах не подрывается. Другое дело в климаксных сериях – здесь чистая продуктивность снижается, и в принципе становится константой. В этом случае очень важно знать величину этой константы, с тем, чтобы четко представлять себе ту величину чистой продукции, которую можно изъять из системы, сохранив ее способность к самовозабновлению. Например, вырубку лесов надо вести на локальных участках с оставлением части территории с коренными типами пород. Это сократить время восстановления фитоценозов, так как сукцессионные серии сократятся до нескольких десятилетий. Сплошная рубка приведет к разрушению всей экосистемы, в том числе эдафической ее части. Восстановление лишь почв потребует тысячелетия. Более того, сукцессионная серия может пойти и не по пути формирования прежнего лесного сообщества, а по пути формирования пустыни и болот или других малопродуктивных экосистем. Таким образом, сообщество не может одновременно быть высокостабильным и давать большой выход чистой продукции, который можно было бы изъять без вреда для самого биоценоза. Развитие растительного покрова никогда не останавливается. Он меняется в силу своего взаимодействия со средой (процессы сингенеза и эндоэкогенеза). Меняя среду, фитоценоз меняется сам. Изменения в составе фитоценоза происходят непрерывно, быстро или постепенно. В ходе этого непрерывного развития неизбежно приходится выделять стадии, поскольку определенная группа видов, доминирующая в данный момент, уступает доминирование другой группе видов, а влияние первой группы уменьшается. Если в силу каких–либо причин растительный покров внезапно нарушен, он восстанавливается относительно быстрыми темпами, но если таких резких нарушений нет, изменения растительного покрова протекают более медленно. Эти изменения происходят даже в тех случаях, когда климатические условия относительно постоянны. При этом фитоценоз изменяется за счет микроэволюции образующих его видов, т. е. путем все более глубоких приспособлений этих видов к условиям фитосреды и друг к другу. Темпы сукцессий неодинаковы. На участке, лишенном растительного покрова, первые стадии сменяют друг друга через год несколько лет, а далее процесс смен замедляется и более позднее стадии восстановления или формирования растительного покрова занимают десятилетия, а затем и столетия. Сложившиеся фитоценозы сменяют друг друга уже в течение тысяч, десятков и сотен тысяч лет. Наконец, продолжительность филоценогенетических смен соразмерна с продолжительностью геологических периодов. Изменения фитосреды, вначале благоприятные для фитоценоза, затем могут стать для него неблагоприятными, что приводит к смене одного фитоценоза другим. Процесс создания фитоценозом неблагоприятных условий для своего существования на первых стадиях развития идет быстрее, при этом обычно быстрее изменяются и условия среды. Все это влечет за собой более скорую смену одного фитоценоза другим; на более поздних стадиях развития фитоценоза эти процессы идут медленно и соответственно более медленно идут смены стадий сукцессии. Вымирания видов под влиянием прямого неблагоприятного действия фитосреды, как правило, не наблюдается; постепенно среда становится благоприятнее для видов, которые имеют в фитоценозе подчиненное значение, и менее благоприятной для видов, господствующих на данной стадии в фитоценозе. В результате первые получают преимущество в борьбе за существование со вторыми и постепенно вытесняют их. Растения, которые при смене фитоценозов остаются в новых фитоценозах, приспосабливаются к новым условиям фитосреды и образуют новые биотипы и экотипы. Таким образом, в процессе смены фитоценозов происходит та микроэволюция, о которой говорилось ранее. В процессе сукцессий возникают биогеоценозы, в наибольшей степени соответствующие условиям среды, как климатическим, так и эдафическим, а также состоящие из «подогнанных» к совместному обитанию видов со свойственным этому ценозу фитоклиматом и гидрологическим режимом. Эта завершающая стадия сукцессии – климакс. Вполне понятно, что в силу своей «пригнанности» к условиям обитания климаксовая стадия гораздо более устойчива, чем начальная и переходные стадии данной серии. Тем не менее, и климакс продолжает изменяться, хотя значительно более медленно, чем остальные стадии. Эти изменения объясняются тем, что никогда не прекращаются изменения внешних условий, вызывающие гологенез, и соответствующая перестройка фитоценоза. Климакс, по мнению основателя теории климакса Клементса, соответствует формации в понимании американских авторов, т. е. зональному типу, или в некоторых случаях подтипу растительности в понимании российских исследователей. Так, формацией следует считать всю зональную растительность тундровой зоны или зональную растительность подзоны хвойных лесов, или зональную растительность подзоны широколиственных лесов. Климакс постоянен до тех пор, пока не изменится климатическая обстановка. Стабильность климакса может быть нарушена только человеком или изменением климата. По представлениям Клементса и его сторонников, при изменении климата на большей части территории климакс не сохраняется, и лишь в благоприятных эдафических условиях остаются реликты прежнего климакса, так называемые постклимаксы . Таким образом, представления Клементса могут быть сведены к тому, что без изменения климата или без воздействия человека зональная растительность не может смениться растительностью, свойственной другой зоне, например, степная растительность не может смениться лесной без изменения климата. Это представление получило название теории моноклимакса , единственного климакса. Согласно этим представлениям, каждая зона имеет лишь один климакс, к которому стремятся все серии сукцессий, в каких бы условиях они ни происходили. Таким образом, по Клементсу, и болота, и поемные луга и растительность склонов стремятся и в конце концов приходят к одному климаксу. Сторонники поликлимакса для всех местообитаний различают сообщества, находящиеся в условиях относительного соответствия местообитанию (климаксы), и сообщества, быстро изменяющиеся (стадии серии, или серийные сообщества). Согласно этим воззрениям, каждое местообитание имеет свой климакс. Этим термином можно и следует пользоваться, понимая под климаксовыми такие сообщества, которые находятся в относительном соответствии с условиями существования и потому изменяются весьма медленно, значительно медленнее, чем сообщества, представляющие собой стадии сукцессионных серий. Следует отметить, что в настоящее время сторонников теории моноклимакса по существу нет. Очень близка к теории климакса теория подвижного равновесия растительного покрова, которую в 20–е годы в России развивали А. А. Еленкин и Л. Г. Раменский. Сочувственно относились к ней и некоторые другие исследователи. За рубежом ее развивали Р. Cepнандep (Sernander, Швеция), А. Тенсли (Tansley, Англия), Каулс (Кawlеs, США) и др. Согласно этой теории, все сообщества можно разделить на три категории: равновесные, равновесно–сменные и нарушенные (Алехин, 1928). Равновесные сообщества характеризуются тем, что их строение и состав колеблются около какой–то средней точки, представляющей как бы типичное состояние растительного покрова. К их числу относится большая часть фитоценозов. Равновесно–сменные сообщества отличаются от равновесных тем, что сама точка равновесия постепенно смещается в каком–либо направлении, и колебание особенностей сообщества совершается вокруг этой движущейся точки. Примером равновесно–сменных сообществ могут быть фитоценозы поймы, где одни участки все время меняются в сторону высыхания, другие – в сторону заболачивания. Нарушенными сообществами называют такие, которые из–за резкой смены условий выведены из состояния равновесия. Эти сообщества стремятся вернуться к тому состоянию равновесия, из которого они были выведены внешней по отношению к ним силой. Таковы сообщества, возникающие при распашке, порубках, пожарах и т. д. Сторонники этой теории, как и некоторые сторонники теории климакса, все развитие сообщества объясняют воздействием внешних факторов. При этом из их поля зрения полностью выпадает самодвижение, развитие фитоценоза в результате действия внутренних причин. Таким образом, теория подвижного равновесия весьма близка к теории климакса в ее первоначальном варианте, когда полагали, что климакс не меняется без изменения внешних условий, и отличается от нее не принципиально, а лишь постановкой вопроса в другой плоскости. Равновесные сообщества соответствуют климаксу (поликлимаксу), но сторонники теории подвижного равновесия подчеркивают колебательные изменения этих сообществ вслед за колебаниями условий среды, а сторонники климакса – неподвижность этих сообществ без изменения условий среды. В свое время теория подвижного равновесия в применении к растительным сообществам сыграла до некоторой степени положительную роль, так как заставила внимательнее присмотреться к погодичным, флюктуационным изменениям сообществ в зависимости от условий погоды разных лет, к влиянию на фитоценозы медленных изменений среды (например, в поймах рек). Однако решала она эти вопросы неправильно, с механистических позиций, недооценивая явления самодвижения, саморазвития фитоценозов. Смена одного сообщества другим совершается в результате изменения конкурентных отношений между видами, образующими сообщество. При сукцессиях, за исключением катастрофических смен, происходящих, например, под влиянием наводнений, пожаров, смена одного сообщества другим идет постепенно: часть видов первого сообщества исчезает быстрее, часть – медленнее, часть остается в новом сообществе, нередко более или менее резко меняя свое фитоценотическое значение. При этом растения одного фитоценоза попадают в другой, конкурируют с компонентами последнего, вытесняют их, занимая их место. При успешном развитии этого процесса количество «наступающих» особей и видов все увеличивается, меняется и структура подвергающегося «агрессии» фитоценоза под влиянием изменения видового состава и соотношения видов, а также изменений среды, и в конечном итоге первый фитоценоз вытесняется вторым. Именно изменение не только видового состава, но и структуры (и сезонного хода развития) фитоценоза позволяет утверждать, что здесь имеет место не только борьба отдельных видов растений, но и фитоценозов в целом, представляющих определенные единства. Эта борьба фитоценозов осуществляется в конкретных условиях и нельзя считать один из фитоценозов во всех случаях более сильным, а другой более слабым. Сила и слабость фитоценозов зависят от экологических условий. Методология изучения вековых смен 1. Многочисленные вековые смены растительного покрова, происходившие в различных районах суши, могут быть до известной степени выяснены с помощью спорово–пыльцевого анализа, широко вошедшего в практику историко–фитоценологических исследований. 2. Некоторое представление о прежнем растительном покрове дают и так называемые реликтовые фитоценозы, сохранившиеся в том или ином районе от прошлых геологических периодов. ПРИМЕРЫ. Так, степные острова в долинах рек Амги (приток Алдана), Лены, Вилюя, Яны и по их склонам или в районе так называемой Кунгурской островной лесостепи (Пермская область) свидетельствуют о том, что степь ранее (очевидно, во время так называемого ксеротермического пустынно–степного периода) была здесь широко распространена. Торфяные сфагновые болота, встречающиеся в подзоне южных степей (Кустанайская область, Казахская ССР), молоды, но они возникли в так называемом субатлантическом периоде, когда климат был значительно более влажным, чем в настоящее время. Сохранение таких реликтовых фитоценозов, чуждых общей климатической обстановке района (степи среди тайги, сфагновыe болота среди южных степей), связано с местными условиями рельефа, почв, вод, создающими возможность сохранения здесь этих фитоценозов. От вековых смен следует отличать возникновение и эволюцию различных типов растительности. Отличие эволюции экосистем от сукцессий заключается в том, что в ходе эволюции появляются новые комбинации видов и вырабатываются новые механизмы их сосуществования. Итогом природной эволюции является разнообразие экосистем, которое было рассмотрено в главе 11. В отличие от организмов экосистемы и их биоты как целостности не эволюционируют. Эволюция экосистем протекает как сеткообразный процесс, который складывается из более или менее независимой эволюции видов, входящих в их состав (Уиттекер, 1980). Для организмов одного трофического уровня главным механизмом эволюции является диверсификация, т.е. усиление несходства видов – эволюция не «к», а «от», что позволяет видам занимать разные экологические ниши и устойчиво сосуществовать в сообществе. Принцип разделения экологических ниш смягчает конкуренцию и может дополняться уже рассмотренными механизмами взаимного (как в семейных группах животных) или одностороннего благоприятствования (как у растений–нянь и их подопечных). Однако диверсификация – это не единственный механизм эволюции организмов одного трофического уровня. В ходе эволюции возможна и унификация экологических характеристик видов. В этом случае, обладая равными конкурентными способностями, виды могут сосуществовать в одном сообществе благодаря влиянию хищников и паразитов, которые ослабляют конкурирующие особи. Кроме того, такие виды могут занимать одну и ту же нишу в разных местах сообщества или поочередно в одном месте. Соотношение диверсификации и унификации, видимо, нетождественно в разных группах организмов. У взаимодействующих организмов разных трофических уровней в ходе эволюции может формироваться широчайшая гамма коадаптаций: от различных форм мутуализма (облигатного или протокооперации), аменсализма, комменсализма до приспособлений, смягчающих антагонистические отношения (между растениями и фитофагами, хищниками и жертвами, хозяевами и паразитами). Коадаптации отношений «растение – фитофаг» и «хищник – жертва» часто имеют диффузный (коллективный) характер: приспосабливаются друг к другу не отдельные виды (вид А– вид Б), а целые гильдии («команды»). Например в саванне приспосабливаются друг к другу «команды» трав и травоядных, древесных растений и веткоядных. Разумеется, приспособление в этом случае означает не взаимопомощь, а снижение интенсивности антагонистических отношений. Вследствие диффузной коадаптации в основе «триплетов» из организмов трех трофических уровней («растение – фитофаг – зоофаг», «фитофаг – хищник первого порядка – хищник второго порядка», «хищник первого порядка – хищник второго порядка – паразит») лежат не пищевые цепи, а пищевые сети, в которых виды могут замещать друг друга. В этом проявляется уже упомянутый принцип: количество функциональных ролей много меньше, чем число их потенциальных исполнителей. В то же время сеткообразность процесса эволюции экосистем не исключает возможности возникновения прочных и однозначных связей, в первую очередь в парах «хозяин – паразит» или при мутуалистических отношениях. Природная эволюция экосистем протекает в масштабе тысячелетий, в настоящее время она подавлена антропогенной эволюцией, связанной с деятельностью человека. Биологическое время антропогенной эволюции имеет масштаб десятилетий и столетий. Антропогенная эволюция экосистем разделяется на два больших класса (по типу процессов): целенаправленная и стихийная. В первом случае человек формирует новые типы искусственных экосистем. Результатом этой эволюции являются все агроэкосистемы, садово–парковые ансамбли, морские огороды бурых водорослей, фермы устриц и т.д. Однако к «плановой» эволюции всегда добавляются «неплановые» процессы – происходит внедрение спонтанных видов, например сорных видов растений и насекомых–фитофагов в агроценозы. Человек стремится подавить эти «неплановые» процессы, но это оказывается практически невозможным. Стихийная антропогенная эволюция экосистем играет большую роль, чем целенаправленная. Она более разнообразна и, как правило, имеет регрессивный характер: ведет к снижению биологического разнообразия, а иногда и продуктивности. Основу стихийной антропогенной эволюции составляет появление в экосистемах видов, непреднамеренно (реже преднамеренно) занесенных человеком из других районов. Масштаб этого процесса столь велик, что принял характер «великого переселения» и «гомогенизации» биосферы под влиянием человека (Lodge, 1993). Заносные виды называются адвентивными (Kornas, 1978, 1990), а процесс внедрения (инвазии) адвентивных видов в экосистемы – адвентивизацией. Причиной расселения адвентивных видов является антропогенное нарушение процессов саморегуляции экосистем при отсутствии видов–антагонистов (Элтон, 1960), как у североамериканской опунции в Австралии и амазонского водяного гиацинта в Африке и Азии, или, напротив, при появлении вида–патогена, к которому у местного вида, ставшего его хозяином, нет иммунитета, как в историях с гибелью Castanea dentata и нарушением африканских саванн вирусом коровьей чумы (см. 8.5). «Экологические взрывы» вызывает занос видов, которые оказываются ключевыми. Чаще такие «взрывы» вовсе не происходят, так как адвентивный вид вообще не вытесняет аборигенные виды из сообщества или если вытесняет, то берет на себя выполнение функциональной роли вытесненного вида. В процессе антропогенной эволюции могут усиливаться и некоторые виды местной флоры и фауны, которые оказались преадаптированными к режиму возрастающих антропогенных нагрузок. В прошлом они были связаны с местами локальных естественных нарушений – горных селей, пороев, вытаптываемых участков экосистем у водопоев, лежбищ крупных фитофагов, таких как зубры или бизоны, и т.д. Результатом антропогенной эволюции экосистем, кроме того, является: – уничтожение видов или снижение их генетического разнообразия (число страниц в Красных книгах во всех странах год от года увеличивается); – смещение границ природных зон – развитие процесса опустынивания в степной зоне, вытеснение травяной растительностью лесов у южной границы их распространения; – возникновение новых экосистем, устойчивых к влиянию человека (например экосистем сбитых пастбищ с обедненным видовым богатством); – формирование новых сообществ на антропогенных субстратах при их естественном зарастании или рекультивации. Однако основу антропогенной эволюции сегодня, безусловно, составляет процесс расселения заносных видов, называемый адвентивизацией. Изучение динамики растительного покрова, различных смен – как кратковременных, так и вековых, позволяет человеку овладеть этим процессом и направить развитие растительного покрова по тому пути, который наиболее целесообразен.
|